

Часть II
РЕФЛЕКС И РЕФЛЕКТОРНАЯ ДУГА
Нервная система в процессе эволюции формировалась как система быстрого реагирования и адаптации к меняющимся условиям внешней среды. Раздражение определенной модальности (свет, звук, механическое воздействие, температура и т.д.) вызывает возбуждение в особых воспринимающих структурах – рецепторах.
Это возбуждение в виде залпа (группы) импульсов передается в центральную нервную систему (ЦНС), перерабатывается и расшифровывается клетками ЦНС. В результате переработки принимается решение, и сигнал-команда в виде импульсов направляется к соответствующему рабочему или исполнительному органу – мышце (гладкой и скелетной) или железистым клеткам.
Структура, с помощью которой осуществляется ответная реакция организма на раздражение из внешней или внутренней среды, называется рефлекторной дугой. А сам ответ – рефлексом. Представления об этом процессе связаны с именем Р. Декарта (XVIII в.). Типичная рефлекторная дуга состоит из 5 или 6 блоков (нарисовать схему).
Рефлекс – основной механизм деятельности нервной системы. Все функции головного и спинного мозга по сути своей – рефлекторны (см. Сеченов И.М., М. 1952).
Простейшая рефлекторная дуга состоит из 2 нейронов: чувствительного (афферентного) и исполнительного – мотонейрона (эфферентного). Между нейронами в цепи всего один синапс. Это моносинаптическая дуга. Если нейронов больше (имеются вставочные или интернейроны), дуга будет дисинаптической или полисинаптической. Рефлексы могут быть врожденными (с соответствующей дугой) и приобретенными в течение онтогенеза.
С развитием рефлекторной теории стало ясно, что нужно ответить на целый ряд вопросов, чтобы понять, как в реальных условиях организм реагирует на множество раздражений, и его мозг (ЦНС) не путается в многочисленных сигналах. Например, во время лекции студент одновременно слушает, смотрит, пишет, и в то же время мозг обеспечивает процессы дыхания, регулирует кровообращение, степень напряжения разных мышц, участвующих в поддержании позы и т.д.
Таким образом, мозг и его части должны одновременно отвечать на самые разные раздражения и адресовать командные сигналы к совершенно разным исполнительным приборам. Соответствующие части мозга должны, как минимум, не мешать друг другу при выполнении своих функций.
НЕРВНЫЙ ЦЕНТР –
ОСНОВНАЯ ФУНКЦИОНАЛЬНАЯ ЕДИНИЦА ЦНС
Часть мозга, которая выполняет какую-либо определенную функцию, называется нервным центром. А.А. Ухтомский, который ввел это определение в научный оборот, назвал нервным центром совокупность нейронов, выполняющую одну общую функцию или являющуюся центром одного рефлекса. Весь мозг состоит из большого количества нервных центров.
Нервный центр – понятие, прежде всего, функциональное. Это означает, что соответствующие нейроны могут располагаться в разных частях ЦНС. Например, клетки, регулирующие дыхание, есть в продолговатом мозгу (собственно "дыхательный центр"), в варолиевом мосту, в промежуточном мозгу и в спинном. А если нужно произвольно изменить характер дыхания, то не обойтись без клеток коры головного мозга.
На практике бывает трудно определить, относятся ли какие-то структуры к соответствующему нервному центру или нет.
А.А. Ухтомский предложил для решения этой задачи использовать критерии, заимствованные из математики. Это критерии 1) необходимого и 2) достаточного. В соответствии с первым, к нервному центру нужно относить структуры, без которых функция или рефлекс вообще не реализуются. Так, дыхательным центром нужно называть соответствующие участки продолговатого мозга, в которых генерируется ритм дыхательных движений. Но у человека этот ритм дыхательным мышцам передается через нейроны спинного мозга. Поэтому, в строгом соответствии с критерием необходимого, соответствующие участки спинного мозга нужно также относить к дыхательному центру. В функциональном отношении эти две структуры действуют как единое целое.
Главная роль в механизме генерирования ритма дыхательных движений принадлежит структурам продолговатого мозга. Остальные структуры мозга могут менять характер дыхания, и без них дыхание не будет адекватно условиям существования организма. Каждый из таких мини-центров выполняет свою часть общей функции – управления дыханием. Работа каждого из них подчиняется общей задаче всего дыхательного ансамбля, согласуется друг с другом.
Согласованная деятельность нервных центров, направленная на решение общей задачи, называется координацией.
Когда говорят о мышечной или какой-то иной координации, то речь всегда идет о согласованной деятельности соответствующих нервных центров. Для обеспечения координации используются различные физиологические механизмы.
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Прежде всего, нужно учитывать, что нервный центр – это группа клеток, которые связаны многочисленными синаптическими связями. Одна нервная клетка может иметь до 100000 синапсов на своей поверхности. Кроме того, отростки многих нейронов ЦНС лишены миелиновой оболочки (Шванновской), характерной для периферических нейронов и для нервных волокон, связывающих разные отделы ЦНС. Благодаря этим обстоятельствам нервные центры обладают и свойствами отдельных нейронов, и свойствами собственно центров, в которых нервные клетки связаны многочисленными синаптическими связями.
Благодаря синапсам возбуждение, проходя через нервный центр, замедляется, и сигналы идут только в одном направлении – от пресинаптических структур к постсинаптическим, то есть от аксонов к дендритам или телу последующей клетки.
Импульсы, которые приходят к центру, суммируются. Способностью к суммации (интеграции) возбуждения и торможения обладает и отдельная клетка. Различают суммацию пространственную и временную. Пространственная суммация – сложение сигналов, которые идут от разных источников или от одного источника (1-ой клетки), имеющего много синаптических контактов за счет веточек собственного аксона (схема).
Временная суммация – сложение сигналов, которые идут по одному пути с определенными интервалами времени (схема) (нервная клетка может возбуждаться только при включении нескольких синапсов одновременно).
В результате суммации (как временной, так и пространственной) в зависимости от того, каких сигналов больше, то есть тормозных или возбуждающих, нервных центр снижает свою активность или усиливает ее.
Ответная реакция нервного ансамбля зависит и от того, как располагаются синаптические связи в пределах этого центра. Если эти связи таковы, что одна клетка образует много синаптических связей с другими нейронами, то ее возбуждение благодаря такой дивергенции будет активизировать сразу много клеток (схема). То есть возбуждение будет иррадиировать в пределах центра или от него к другим центрам. Значит, информация быстро распространится в пределах центра, будет передаваться большему числу структур. Такой принцип функциональной организации характерен для симпатической нервной системы. Благодаря нему возбуждение охватывает большое количество исполнительных структур вегетативного обеспечения функций, мобилизует ресурсы, необходимые для адаптации, например, во время стресса.
Но организация синаптических связей в пределах центра или между ними может быть по конвергентному принципу: много нервных клеток, афферентных и промежуточных, связаны с небольшим количеством мотонейронов (схема). В результате возбуждение большого количества афферентных клеток активизирует относительно немного мотонейронов.
Так как каждый мотонейрон связан с несколькими афферентными или промежуточными клетками одновременно, суммарная реакция мотонейронов на раздражение афферентов может быть меньше, слабее, чем можно ожидать при раздражении частей афферентов.
Сложные синаптические связи между клетками нервного центра приводят к тому, что клетки образуют различные сети, кольцевые структуры, в которых может долго без затухания циркулировать возбуждение. Такое возбуждение называется тоническим. Если такой центр управляет мышцами, то мышцы будут находиться в состоянии небольшого напряжения – тонуса. Тонус, по-видимому, помогает поддерживать постоянное рабочее состояние. При необходимости, тонический режим можно легко усилить или, наоборот, затормозить.
Для некоторых больших структурно-функциональных образований мозга характерна постоянная тоническая активность. Таким очень важным образованием является ретикулярная формация (РФ) ствола мозга. Она оказывает возбуждающее влияние на кору больших полушарий, исполняет роль своеобразной энергетической станции головного мозга. Благодаря этому кора больших полушарий находится в состоянии тонуса, необходимого для рабочей деятельности, если уровень возбуждения РФ снижается, активность коры также падает. Это характерно, в частности, для состояния сна.
Иногда тоническая активность нервных центров носит правильный, упорядоченный характер. Это характерно, например, для дыхательного центра. Такой вид тонуса отвечает за определенный функциональный ритм, а также может служить своего рода, биологическими часами. Это может быть очень важной функцией нервных центров.
Биологические часы отсчитывают суточные ритмы (например, сон – бодрствование), а также ритмы с более длинным (недели, месяцы и более) периодом. Исходя из общих соображений, можно предположить, что с помощью таких "часов" можно отсчитывать время последовательного включения мышц при выполнении сложно-координированных движений.
Однако важно отметить, что тонус нервных центров во многих случаях может меняться под действием раздражений, поступающих от рецепторов. Известно, что ритм дыхательного центра продолговатого мозга меняется при изменении концентрации СО2 и О2 в крови. Пределы изменения ритма, по-видимому, определяются таким важным свойством нервного центра, как лабильность.
В ряде случаев нервному центру можно навязать какой-то новый ритм работы. Это можно легко обнаружить при раздражении некоторых нервных центров низших животных (например, лимфатического центра амфибий). Относительно легко можно менять ритм работы, многих двигательных центров человека. Именно поэтому в процессе тренировок можно обучить нервные центры работать чаще, в более высоком ритме. Правда, увеличения ритма не очень велики.
КООРДИНАЦИЯ НЕРВНЫХ ЦЕНТРОВ.
ДОМИНАНТА
Тонус центра, то есть уровень его возбуждения, может быть очень выраженным. Возбуждение центра может быть очень мощным и преобладать по отношению к другим.
Такой мощный очаг возбуждения называется доминантным. Если есть такой очаг, то и функция, связанная с ним, становится доминирующей, главной в организме. Организм начинает работать, в первую очередь, на обеспечение этой функции. Доминирующий очаг как бы подчиняет себе текущую деятельность организма.
Доминанта обладает способностью тормозить деятельность окружающих нервных центров, что, в свою очередь, облегчает преобладание функции, связанной с доминантой. Это легко можно проверить у человека, поглощенного каким-то делом. Играющий ребенок не слышит зова матери, которая приготовила для него обед. Человек, погруженный в свои мысли, не слышит и не видит автомобиля, который может на него наехать. Кроме того, доминанта способна "перехватывать" возбуждение, вовсе ей не адресованное, и усиливать за счет этого уровень своего возбуждения. Это можно обнаружить, если голодного щенка попытаться оттащить за хвост от блюдечка с молоком. Вместо того чтобы огрызнуться на постороннее мешающее раздражение, щенок быстрее лакает молоко. Если плачущего ребенка начать утешать, гладить по головке, то результат, как правило, обратный – он плачет еще сильнее.
Анализ ситуации с плачущим ребенком подсказывает способ подавления (ликвидации) доминанты. Если внимание ребенка переключить на что-то интересное, то он перестает плакать. Причина этого заключается в том, что интересное явление, дело, предмет вызывают к жизни другой доминантный очаг. А эта доминанта, если она мощнее первой, тормозит ее.
После затухания доминанты остается след в виде повышенной возбудимости. Этот след – физиологическая основа памяти. Таким образом, выстраивается цепочка: 1) доминанта, которая обеспечивает нужный уровень внимания, отсекая лишнее возбуждение; 2) концентрация внимания, которая способствует формированию следа события, а, значит, – 3) памяти.
А почему возникает доминанта? Ее появление связано с потребностями организма. Проще всего анализировать соотношения между биологическими потребностями и доминантой.
К основным биологическим потребностям относятся пищевые, половые, информационные и некоторые другие. Нейрофизиологической основой проявления этих потребностей является работа различных подкорковых структур. Одной из важнейших среди них является ретикулярная формация (схема). Ретикулярная формация (РФ) занимает центральную часть ствола головного мозга. За счет восходящих путей оказывает мощное возбуждающее влияние на кору больших полушарий и отдельные ее части. РФ, как отмечалось ранее, – это своеобразная энергосиловая станция мозга. Если отсечь ее влияние (с помощью перерезки) на кору больших полушарий, то животное в эксперименте теряет всякую активность и находится в сноподобном состоянии. Сходное состояние обнаруживается и у человека при поражении РФ вирусом или каким-либо другим фактором.
Если человек хочет есть, то это означает определенную последовательность событий. В крови понижается концентрация глюкозы. Сигнал об этом соответствующие хеморецепторы передают в головной мозг. Центры голода располагаются в РФ. Они возбуждаются. Если возбуждение сильное, центры могут стать доминантными. Подкорковая доминанта возбуждает соответствующие участки коры. Человек ощущает чувство голода. Соответствующий доминантный очаг заставляет так организовать поведение человека, чтобы удовлетворить это чувство голода. Если в результате этого удалось поесть, то меняется активность рецепторов глюкозы, падает возбуждение в нервных центрах. Доминанта исчезает.
Таким образом, доминанта так организует поведение, чтобы ликвидировать саму себя в результате удовлетворения соответствующей потребности.
Поскольку доминанта подчиняет себе текущую деятельность организма, выполняет системообразующую функцию, то она определяет выбор реакции, рефлекса. Из множества вариантов мозг выбирает тот, который направлен на удовлетворение главной в данный момент потребности. Поэтому управление доминантой – это управление соответствующими потребностями. Если научиться доминантой управлять произвольно, то можно произвольно же и регулировать потребностно-мотивационную сферу человека, отсекать ненужное волнение, посторонние мысли, повышать интерес к делу и, как правило, улучшать качество запоминания, глубину памяти и т.д.
Учение о доминанте, разработанное замечательным отечественным физиологом А.А.Ухтомским, позволяет понять, как из множества афферентных сигналов, запускающих разные рефлекторные дуги, мозг выбирает что-то немногое, наиболее важное для него в данный момент времени, соответствующее его потребностям. Из возможных рефлексов работает только ограниченное их количество, а проявление остальных с помощью доминанты тормозится.
Иными словами, доминанта – важнейший механизм координации (согласованной деятельности) функций, в основе которой лежит координация нервных центров.
ОБЩИЙ КОНЕЧНЫЙ ПУТЬ
Этот механизм лежит в основе ряда других так называемых принципов координации. Например, общего конечного пути, открытого, как принято считать, Ч. Шеррингтоном, английским физиологом, нобелевским лауреатом. Правда, сформулирован этот принцип фактически И.М. Сеченовым в его гениальной брошюре "Рефлексы головного мозга". Сущность этого принципа координации сводится к тому, что огромное количество афферентных структур (чувствительных нейронов) связано с небольшим количеством исполнительных. Иными словами, мозг получает и перерабатывает огромное количество разнообразной сенсорной информации, а следствием этого может явиться сокращение небольшого числа мышц. Например, человек берет в руки авторучку и пишет слово "мама".
Наблюдается такая своеобразная воронка (схема). А как же мозг решает, какие именно исполнители нужно включить?
Выбор определяет доминанта. Выбор этот осуществляется неосознанно (у животных и в ряде случаев у человека) или сознательно. В результате выбора поведение строится так, чтобы удовлетворить соответствующую потребность, а значит уменьшить соответствующее возбуждение.
ИРРАДИАЦИЯ
Прежде чем выключить ненужные нервные центры, мозг должен получить большое количество информации для оценки ситуации. А каждый нервный центр должен "знать", что делают другие. Поэтому информация, которую получит один центр мозга, распространится и к другим нервным центрам. Этот координационный принцип называется иррадиацией.
Благодаря иррадиации, по-видимому, осуществляется своеобразный перебор всех возможных вариантов организации функциональной системы, объединение различных органов, которое решает какую-то задачу, добивается конкретного полезного результата. А доминанта оказывает определяющее влияние на выбор главного нервного центра, которому подчиняются остальные.
Доминанта же и вытормаживает "ненужные" нервные центры, используя механизмы постсинаптического или пресинаптического торможения.
ТОРМОЖЕНИЕ В ЦНС
Торможение в ЦНС открыл И.М. Сеченов. При исследовании рефлекса Тюрка он обнаружил, что наложение кристаллика поваренной соли на срез головного мозга (на уровне таламуса) замедляет отдергивание лапки из раствора с кислотой. Причиной этого является тормозное влияние ретикулярной формации на нейроны спинного мозга. Механизм этого торможения пресинаптический.
Открытое И.М. Сеченовым явление – один из важнейших механизмов координации. С помощью торможения выключаются ненужные нервные центры, включаются необходимые, регулируется продолжительность работы различных нервных структур, даже организуется согласованная деятельность целых отделов ЦНС (например, большинство синаптических связей в мозжечке – тормозные).
Торможение, по-видимому, играет очень важную роль в регуляции особых состояний мозга, таких как сон, гипноз. Торможение – ключевой механизм в основе важного координационного принципа – реципрокной иннервации, открытого Ч. Шеррингтоном. Этот принцип лежит в основе управления ритмическими движениями типа ходьбы, бега и т.д. Механизмы торможения (специальные тормозные клетки) обеспечивают периодические включения и выключения центров мышц-антагонистов (схема).
Разнообразные механизмы координации позволяют очень гибко регулировать различные функции и, особенно, движения. В основе движения лежит сократительная деятельность скелетных мышц, которыми управляют соответствующие нервные центры. Движение – это один из компонентов поведения человека, понимание механизмов которого – ключ к пониманию механизмов организации и функционирования ЦНС.
МЕХАНИЗМ И ЭНЕРГЕТИКА МЫШЕЧНОГО
СОКРАЩЕНИЯ
Каждая мышца состоит из множества мышечных клеток (волокон). Каждое такое волокно сокращается, если получает команды в виде импульсов от соответствующего мотонейрона. В каждом мышечном волокне находится множество миофибрилл – тонких пучков сократительных нитей. Эти нити состоят из белков: миозина и актина (схема).
Актиновые нити прикрепляются к поперечным (относительно длины мышечных волокон) перегородкам-пластинкам. Пластинки делят миофибриллы на саркомеры длиной 2,5 мкм. Укорочение мышечного волокна – результат сокращения множества саркомеров. При укорочении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине пучка и саркомера. Сами нити при этом не укорачиваются.
Такое перемещение обеспечивается так называемыми поперечными мостиками. Мостики – это поперечные выступы молекул миозина, которые отходят от нити биполярно. Для того чтобы мостики потянули актиновую нить, их нужно активировать. Активация обеспечивается с помощью ионов кальция. Са2+ снимает блокирующие действия белка тропомиозина, который без Са2+ препятствует прикреплению поперечных мостиков к актиновым нитям. Процесс сокращения требует затрат энергии молекул АТФ. Са2+ находится в продольных трубочках (цистернах), расположенных вдоль миофибрилл. Каждая трубочка располагается под саркомером, между пластинками. Под действием возбуждения Са2+ выходит из этих трубочек и может влиять на работу миофибрилл. Возбуждение с поверхности мышечной мембраны может проникать вглубь волокна, к продольным трубочкам с помощью систем поперечных трубочек (Т-системы), которые обычно окружают каждую миофибриллу на уровне пластинок.
Мышечное сокращение осуществляется в такой последовательности. Нервные импульсы (для возбуждения одной мышечной клетки достаточно одного импульса) через синапс запускают механизм возбуждения мышечного волокна. Возбуждение распространяется вдоль клетки и по Т-системе достигает кальциевых резервуаров. Са2+ выбрасывается из трубочек, снимает блокаду миозиновых мостиков. За счет эластической тяги мостиков актиновые нити втягиваются между миозиновыми. Этот процесс обеспечивается энергией молекул АТФ. Эта энергия освобождается с помощью ферментов, основным из которых является сам миозин.
РЕГУЛЯЦИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ
При одиночном сокращении процесс укорочения заканчивается, Са2+ с помощью специального насоса возвращается в систему каналов, и происходит расслабление мышцы.
Если стимулы поступают с высокой частотой, уровень Са2+ не успевает после каждого сокращения вернуться к норме. Отдельные сокращения при этом сливаются. Наблюдается так называемый тетанус. Частота слияния тем ниже, чем больше длительность одиночного сокращения.
Каждое одиночное сокращение одного мышечного волокна запускается пороговой интенсивностью раздражения. Так называемые быстрые скелетные мышечные волокна подчиняются закону "Все или ничего". Но поскольку мышца состоит из разных по возбудимости мышечных волокон, то ответ целой мышцы будет зависеть от силы раздражения (график).
Такие разные характеристики мышечных волокон позволяют плавно менять величину мышечного напряжения (силу).
Мышечное сокращение целиком зависит от работы соответствующих командных нейронов. Сначала включаются самые возбудимые нервные клетки, затем менее возбудимые. Уставшие клетки могут отключаться. Необходимый уровень напряжения мышцы обеспечивается оставшимися нейронами. Контроль за напряжением ЦНС осуществляет с помощью обратных связей, основу которых представляют проприорецепторы (рецептор в мышцах, сухожилиях и т.д.).
СИЛА МЫШЕЧНОГО СОКРАЩЕНИЯ
Регуляция мышечной силы, однако, в значительной степени – функция ЦНС. Каждая мышца иннервируется мотонейронами. Один мотонейрон связан с определенным количеством мышечных волокон, как правило, от десятков до тысяч клеток. Такой комплекс называется двигательной единицей. Количество элементов в двигательной единице зависит от ее функций. В мышцах, выполняющих мелкие и точные движения, мотонейрон управляет несколькими мышечными волокнами, а в камбаловидной мышце двигательная единица включает в себя более 1000 мышечных клеток. Так как мышечные волокна сокращаются по закону "Все или ничего", двигательная единица или работает вся или вся отдыхает.
Двигательные единицы обладают разной возбудимостью. Поэтому они могут по-разному включаться в процесс сокращения и обеспечивать разную величину мышечного напряжения. Кроме того, развиваемая сила зависит от частоты стимуляции. При высокой частоте разрядов мотонейронов одиночные сокращения сливаются, что приводит к суммации силы.
Во время полного, так называемого гладкого тетануса, сила примерно в 2 раза больше, чем при неполном (зубчатом) тетанусе, который развивается при низких частотах стимуляции.
Комбинация этих механизмов позволяет ЦНС при необходимости увеличивать мышечную силу до максимума. Частота раздражения, которая вызывает максимальное мышечное сокращение, называется оптимумом. В этих условиях, по-видимому, в процесс сокращения вовлечены все мышечные волокна. Дальнейшее увеличение частоты или силы раздражения уже не может вызвать прибавку силы, и она может даже снижаться (объяснить, почему?).
Величина мышечной силы зависит от толщины мышцы (физиологического поперечника), точнее, от количества миофибрилл в мышечных волокнах. По этой причине изометрические упражнения дают большую прибавку в силе, так как стимулируют синтез сократительных белков. Изометрические упражнения – мышечная активность, при которой не меняется длина мышцы, но увеличивается мышечное напряжение. Движения с изменением длины относятся к группе динамических. Мышечное сокращение, когда длина мышцы уменьшается с увеличением ее силы, называют ауксотоническим.
В основе мышечного сокращения лежит взаимодействие нитей актина и миозина. Если взаимодействие (реакция) между ними ослабевает, то ослабевает мышечная сила. Это ослабление проявляет себя либо при сильном укорочении саркомера (сократительные нити начинают мешать друг другу), либо при очень большом растягивании capкомера (актиновые и миозиновые нити не перекрываются) (привести примеры).
СИЛА И СКОРОСТЬ
Сила зависит и от скорости движения. Во время быстрого укорочения мышца развивает меньшую сократительную силу, чем при медленном сокращении (это вытекает из закона Холла о связи нагрузки со скоростью). Поэтому тяжелый вес можно поднять достаточно медленно. Однако и в этом случае скоростью движения можно управлять произвольно. Если мобилизовать много мышечных волокон (в идеале – все), относительная нагрузка на каждое активное мышечное волокно получается меньше, скорость их сокращения будет больше, чем в случае активации небольшого количества мышечных волокон. Таким образом, вовлечение дополнительного количества двигательных единиц при одной и той же нагрузке позволяет увеличить скорость движения.
Сила, которую развивают несколько мышц, участвующих в каком-то движении, не равна арифметической сумме напряжений каждой из мышц. Существенную роль играют расположение мышц, характер суставов, участвующих в движении, и управления ЦНС, то есть координационные механизмы.
Роль ЦНС возрастает при сложных реакциях, которые опираются на полисинаптические рефлекторные цепи. Чем лучше функционирует такая цепь, т.е. чем быстрее по ней пробегает возбуждение, тем меньше время реакции.
Важную роль играют функциональные свойства моторных единиц. У человека быстрые мышечные сокращения обеспечиваются "быстрыми" моторными единицами, в состав которых входят мышечные волокна, которые быстро сокращаются, но не могут долго работать из-за особенностей метаболизма. Работа таких структур обеспечивает скорость движения. Если соответствующие нервные центры могут функционировать в высоком ритме, на выходе возможна высокая скорость движений.
Для длительной работы необходимы мышечные структуры с экономными механизмами энергообеспечения, высоким коэффициентом полезного действия, использование энергии и совершенные координационные механизмы. Задача ЦНС – соединить уровень мышечного сокращения с соответствующими вегетативными системами обеспечения в единое целое.
ФИЗИОЛОГИЯ МЫШЕЧНЫХ РЕЦЕПТОРОВ
Очень важную роль в координации мышечной активности играют обратные связи. С их помощью обеспечивается два вида двигательных функций: поддержание положения (позы) и собственно движения. Нейрофизиологические механизмы организации двигательных функций довольно хорошо изучены. Их понимание позволяет лучше разобраться в рефлекторных механизмах регуляции всех функций. Любой рефлекторный ответ начинается с включения соответствующих рецепторов. Именно с их помощью соответствующие нервные центры получают информацию о сокращении мышц, о положении суставов, о характере мышечного движения.
В каждой мышце обнаруживаются волокна, которые тоньше и короче других. Такие волоконца располагаются небольшими скоплениями, окруженными соединительнотканной оболочкой. Эти структуры называются мышечными веретенами, так как имеют веретенообразную форму. Мышечные волокна в них называют интрафузальными, в отличие от экстрафузальных, из которых состоят рабочая мышца.
Существует два типа интрафузальных волокон, отличающихся по размеру и расположению многочисленных клеточных ядер. Волокна своими концами прикрепляются к экстрафузальным волокнам с помощью как бы маленьких сухожильных полосок соединительной ткани длиной 0,5-1,0 мм.
Каждое веретено связано с одним дендритом. Веточки дендрита связаны с каждым интрафузальным волоконцем (схема). Каждая веточка обматывается вокруг средней части интрафузального волоконца в виде спирали – аннулоспирального окончания. Есть еще афферентная иннервация с помощью других нервных волокон. Они связаны с периферическими частями интрафузальных волокон.
Кроме того, интрафузальные мышечные волокна иннервируются так называемыми γ-мотонейронами. Один γ-мотонейрон управляет сокращениями нескольких мышечных веретен.
Число мышечных веретен в мышце может достигать нескольких сотен, но в расчете на единицу массы веретен больше в мелких мышцах, отвечающих за тонкое движение (привести примеры).
В участках сухожилий, прилегающих к мышцам, располагаются так называемые рецепторы Гольджи, или сухожильные органы.
Мышечные веретена и сухожильные органы при растяжении мышц усиливают свою деятельность. Кроме того, частота импульсации от мышечных веретен зависит и от скорости растяжения (сокращения) мышц, то есть мышечные веретена передают информацию динамического характера и статического.
При изометрическом сокращении активность веретен снижается, а сухожильных – резко возрастает.
Чувствительность мышечных веретен может меняться за счет γ-мотонейронов, которые управляют сокращением интрафузальных волокон. Укороченное интрафузальное волокно начинает передавать информацию о длине экстрафузальных волокон даже при их укорочении. Причем, разные γ-мотонейроны по-разному влияют на статические и динамические характеристики 2-х типов рецепторных волокон. Один тип волокон становится более чувствительным к скорости растяжения, а другой – к определенному уровню растяжения.
Мышечные веретена передают информацию в ЦНС, используя которую мозг очень точно может управлять мышечным сокращением, регулировать тонус мускулатуры.
Например, при растяжении мышцы возбуждаются мышечные веретена. Волокна от них моносинаптическим путем активируют соответствующие мотонейроны, и мышца сокращается, что приводит к уменьшению ее растяжения. Такой механизм работает в поддержании постоянства тонуса, обеспечивающего сохранение позы. Этот механизм способствует сохранению постоянной длины мышцы, автоматически компенсирует изменение нагрузки на мышцу.
Таким образом, рефлекторная дуга, обеспечивающая включение соответствующей мышцы, превращается в рефлекторное кольцо.
Кроме мышечных веретен и сухожильных органов в организации двигательных рефлексов участвуют кожные рецепторы, суставные рецепторы и свободные нервные окончания, расположенные в мышцах.
Соответствующие рефлекторные дуги более сложно устроены, они полисинаптические, и поэтому рефлексы очень пластичны. Раздражение кожных рецепторов, в частности, включает сгибательный рефлекс. Это типичный оборонительный рефлекс, примером которого является рефлекс Тюрка. При этом автоматически тормозятся нейроны разгибателей, и активируются нейроны разгибателей противоположной конечности (что это позволяет сделать?).
Рецепторные образования могут запускать и очень сложные нервные сети. Причем ритм работы нервного ансамбля не зависит от частоты раздражения. Так организована работа локомоторных центров конечностей. Двигательные рецепторы, главным образом, обеспечивают согласованную деятельность этих центров.
Локомоторные центры есть, по-видимому, и у человека. Если раздражать кожу новорожденного, то возбуждение этих центров проявляется в виде шагательного рефлекса новорожденных.
ПОСТРОЕНИЕ ДВИЖЕНИЯ
Таким образом, даже на уровне спинного мозга существуют не только рефлекторные, то есть запускаемые внешними стимулами, но и запрограммированные цепные двигательные акты, способные осуществляться без каких-либо внешних влияний. Такие двигательные программы, как правило, являются врожденными, как, например, дыхание. Подобные программы формируются также в онтогенезе. В этом случае в их образовании участвуют все "этажи" ЦНС: кора больших полушарий, подкорковые структуры, мозжечок, спинной мозг. Каждый уровень при этом решает свои задачи, отвечает за определенные функции.
Спинной мозг – это исполнитель команд, получаемых из высших инстанций. Ствол головного мозга – это, прежде всего, позные двигательные рефлексы. Эти рефлексы включаются, в основном, от рецепторов шеи и вестибулярного аппарата. Основными двигательными центрами ствола являются:1) красное ядро (средний мозг), 2) латеральное вестибулярное ядро Дейтерса, 3) некоторые отделы ретикулярной формации моста и продолговатого мозга. Нейроны этих центров оказывают разное влияние на α-мотонейроны сгибателей и разгибателей. Например, структуры продолговатого мозга возбуждают мотонейроны сгибателей и тормозят мотонейроны разгибателей, а мостовые – наоборот (схема).
Особую роль в регуляции движения и поддержания позы играет мозжечок. Многие движения могут оптимально осуществляться только при его участии. В то же время мозжечок не является жизненно необходимым органом. По-видимому, основное значение мозжечка состоит в дополнении и коррекции деятельности остальных двигательных центров.
Центральная часть мозжечка (промежуточная) участвует в коррекции медленных движений и поз и координации позных и целенаправленных движений. Полушария мозжечка собирают информацию от всех областей коры. Благодаря этому информация о замысле движения превращается в полушариях мозжечка и зубчатом ядре в программу движения, которая посылается в моторную зону коры больших полушарий. Только после этого возможно нормальное движение (схема).
Третий "высший" уровень организации движения представлен двигательной корой и базальными ядрами. Эти структуры имеют особое значение для целенаправленных произвольных движений.
Двигательная кора организована соматотопически, то есть определенные участки коры управляют определенными частями тела. Минимальной функциональной единицей коры является кортикальная колонка, которая включает в себя несколько сотен пирамидных клеток. Такие колонки могут возбуждать или тормозить группу функционально однородных мотонейронов. Исследования показывают, что двигательная колонка управляет несколькими мышцами, действующими на тот или иной сустав. Это означает, что кора управляет не столько мышцами, сколько движениями. Например, она выбирает мышцы, обеспечивающие соответствующие движения.
У человека самым развитым является пирамидный тракт, с помощью которого кора оказывает возбуждающее влияние на мотонейроны сгибателей и тормозное – на мотонейроны разгибателей. Базальные ганглии тесно связаны с моторной корой, участвуя в программированном движении.
Кора больших полушарий (КБП) выполняет не только двигательные функции.
В определенные участки коры приходят сигналы от органов чувств. Эти участки – центры соответствующих анализаторов или сенсорных систем. Есть участки, которые выполняют исключительно "человеческие" функции – анализаторы восприятия слуховых и зрительных речевых образов. Они расположены рядом с корковыми зонами слуха (височная извилина) и зрения (медиальная поверхность затылочной доли). Речевые анализаторы у правшей, как правило, находятся в левом полушарии, у левшей – в правом.
Кроме того, в коре выделяют ассоциативные зоны, которые прямо не связаны с органами чувств или мышцами. Ассоциативные нейроны выполняют интегративно-аналитические функции, благодаря чему формируются целостное поведение, логическое мышление и т.д.
Основные ассоциативные зоны – это теменная доля (задняя и нижняя части), лобная (кроме двигательной части) и лимбическая. Каждая из них особенно важна, соответственно, для построения целостных сенсорных образов и речи, высших двигательных функций, выбора, планирования и запуска поведенческих программ, памяти и эмоциональных компонентов поведения.
Кора работает как единое целое, но левое полушарие у правшей доминирует, в том числе и в двигательной сфере. Оно более тесно связано с речью и словесно-логическим мышлением. Правое полушарие больше связано с чувственной сферой.
Организация движения, таким образом, оказывается многоуровневой, включающей в себя различные компоненты ЦНС и периферических исполнителей. Кроме того, нормальная функция не может осуществляться без вегетативных систем жизнеобеспечения, прежде всего, без систем кровообращения, дыхания, благодаря которым мозг и мышцы во время работы получают кислород и питательные вещества, и выделительной системы, обеспечивающей удаление ненужных и токсических продуктов, образующихся во время работы.
Остальные системы играют основную роль в процессах восстановления (перечислить какие).
И, наконец, очень важную роль в обеспечении высокой работоспособности играют гуморальные регуляторные системы, среди которых особое место занимает система желез внутренней секреции. Нервную регуляцию деятельности органов и систем осуществляет вегетативная нервная система.
ФУНКЦИИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Главная функция вегетативной нервной системы заключаемся в поддержании постоянства внутренней среды (гомеостаза) при различных воздействиях на организм. Таким образом, вегетативная нервная система является важным компонентом рефлекторного системного ответа организма на раздражение. Поэтому вегетативные нервные центры подчиняются тем же законам координации, что и все остальные. Единственное отличие состоит в том, что нейроны вегетативной нервной системы, как правило, более медленно работающие, нежели соматические. Поэтому и центры менее лабильны, чем центры, управляющие скелетной мускулатурой.
Центры вегетативной нервной системы располагаются в грудных и верхних поясничных сегментах спинного мозга (симпатическая н.с.) и в стволе мозга, и крестцовых ceгментах спинного мозга (парасимпатическая н.с). Симпатический отдел и парасимпатический имеет и целый ряд других анатомических отличий (перечислить все отличия!).
Функциональное влияние этих двух отделов часто носит антагонистический характер. Так, раздражение симпатических нервов приводит к увеличению частоты сокращений сердца и ударного объема сердца, ослаблению моторики кишечника, расслаблению желчного пузыря, бронхов, сокращения сфинктеров желудочно-кишечного тракта. Стимуляция парасимпатических нервов (например, блуждающего) вызывает противоположные эффекты.
Однако в большинстве случаев оба отдела действуют согласованно для обеспечения полезного приспособительного результата.
Например, если повышается артериальное давление, возбуждение барорецепторов активирует парасимпатические структуры, регулирующие работу сердца, и одновременно снижает деятельность соответствующих симпатических волокон. Такая функциональная синергия приводит к снижению частоты и силы сердечных сокращений.
Рефлекторные дуги вегетативных рефлексов более сложно устроены. Так, спинальная дуга состоит, как минимум, из 4-х нейронов (нарисовать схему). Кроме того, спинно-мозговой отдел симпатической нервной системы имеет черты сегментарной организации. Эта организация сохраняется для чувствительной и вегетативной иннервации некоторых органов. Вследствие конвергенции афферентов, принадлежащих определенному сегменту на структурах, которые связаны с кожной чувствительностью, заболевания внутренних органов воспринимаются как кожные боли соответствующей локализации. Это так называемые зоны Геда. Например, при заболеваниях желудка болевые ощущения локализуются в левой стороне нижней части живота.
Основной единицей, регулирующей функции организма, управляющей мышцами, является нервный центр. Нервные центры взаимодействуют между собой (координируют) для выполнения различных функций. Механизмы координации соматических и вегетативных функций очень сложны и многоплановы. Но именно эти механизмы позволяют нервным центрам, всему мозгу с удивительным совершенством обеспечивать быструю и точную адаптацию организма к меняющимся условиям внешней и внутренней среды.
Огромная роль в этих механизмах принадлежит синаптическим связям в пределах каждой рефлекторной цепи и между различными нервными структурами.
Любой синапс, как отмечалось ранее, использует вещества-посредники – медиаторы. Медиаторы – это различные химические соединения. Медиатор в центральных нейронах взаимодействует с рецепторами на постсинаптической (иногда на пресинаптической) мембране и запускает изменение внутриклеточного метаболизма с помощью вторичных посредников в случае метаботропных рецепторов. В случае присутствия более быстро работающих ионотропных рецепторов сразу активируется ионный канал.
Вещества, с помощью которых можно изменить состояние того или иного нервного образования, как правило, действуют на синаптический процесс. Если они действуют подобно медиатору, это – агонисты. Если блокируют работу синапса – антагонисты. На пути попадания искусственных агонистов или антагонистов в мозг стоят барьеры – гематоэнцефалический и кишечный (если вещества вводятся через рот – per os).
Рецепторы, на которые действуют медиаторы или их заместители, неоднородны и реагируют неодинаково. Это, по-видимому, обеспечивает более тонкое и точечное влияние на соответствующие клетки. В случаях лекарственного использования тех или иных веществ приходится подбирать нужную концентрацию, что не всегда можно сделать.
АЦЕТИЛХОЛИН
Например, ацетилхолин – медиатор в нервно-мышечных синапсах. Агонистом его является никотин, антагонистом – тубокурарин, который может постепенно выключить все нервно-мышечные синапсы, в том числе и в дыхательных мышцах.
Рецепторы в ЦНС более чувствительны к мускарину (вещество, содержащееся в мухоморах). Его эффекты носят так называемый парасимпатический характер: снижение артериального давления и сердечной активности, повышенное потоотделение, слезотечение, тошнота. Антагонистом мускариновых рецепторов является атропин (токсин белены и дурмана). Эффекты его (периферические) – обратные. Кроме того, наблюдаются центральные эффекты, такие как двигательное и речевое возбуждение, галлюцинации. Последнее означает, что мозговые нейроны, выделяющие ацетилхолин, должны оказывать тонизирующее влияние на определенные участки коры.
Ацетилхолин вырабатывается частью нейронов ретикулярных ядер моста и интернейронами полосатого тела базальных ганглиев, нейронами, участвующими в двигательных системах и ряде других.
НОРАДРЕНАЛИН
Норадреналин – медиатор на периферии (симпатические синапсы) и в ЦНС. В ЦНС норадреналин вырабатывается нейронами голубого пятна моста и межножкового ядра среднего мозга. Аксоны этих нейронов проецируются в спинной и головной мозг, влияя на их состояние. Это, прежде всего, создание определенного уровня активации (тонуса) ЦНС, различных ее участков. Например, норадреналиновые нейроны выключают тормозные интернейроны в моторных центрах и повышают уровень двигательной активности; влияют на выраженность эмоций в стрессовых ситуациях, эмоций, связанных с азартом.
Медиатор необходим для долговременного изменения свойств синапсов в корковых колонках и мозжечке, что позволяет фиксировать новую информацию. Понижение активности норадреналиновой системы может привести к апатии, депрессии, ухудшению памяти.
Рецепторами норадреналина являются α и β, которых много в сосудах, сердце, бронхах. Их активация вызывает разнообразные вегетативные эффекты. Например, действие на α-рецепторы сосудов вызывает их сужение, на β – стимулирует работу сердца, что повышает возможности организма. Аналогично действует агонист норадреналина – эфедрин. Эфедрин оказывает и возбуждающее влияние на мозг, к нему может развиться привыкание.
Инактивация норадреналина осуществляется путем обратного всасывания и разложения с помощью фермента моноаминоксидазы (МАО). Этот фермент разрушает и другие родственные моноамины: дофамин и серотонин.
Норадреналиновая система работает сопряженно с эндокринными механизмами, прежде всего с системой секреции адреналина.
ДОФАМИН
Дофаминовые нейроны находятся в трех местах головного мозга: черной субстанции, покрышке среднего мозга и в ядрах таламуса.
Дофаминовые нейроны черной субстанции связаны с клетками скорлупы и хвостатого тела (полосатое тело), входящими в группу базальных ядер. Функция этих нейронов – поддержание определенного уровня двигательной активности (в частности, необходимого для запуска движения), обеспечение точности выполнения моторных программ, устранение непроизвольных движений.
При разрушении черной субстанции, что характерно для паркинсонизма, затрудняется запуск движений (акинезия), наблюдается патологическое усиление мышечного тонуса и тремора (дрожания пальцев и головы), ряд нарушений эмоционального характера, памяти.
|
В норме
При патологии |
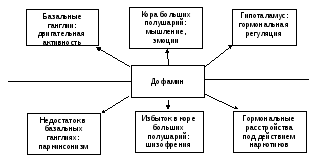
Рис. 1. Эффекты дофаминергической системы в норме и патологии (по Дубынину В.А., 2003)
Основная задача дофаминовой системы – регуляция двигательных эффектов, в том числе тех, которые вплетены в другие реакции, например, эмоции. Дофаминовые нейроны тонко связаны с холинергическими, понижают их активность. Поэтому некоторые симптомы паркинсонизма (тремор) можно ослабить антагонистами ацетилхолина, например, производными атропина.
Дофамин взаимодействует с 5 разными рецепторами. Больше всего так называемых Д1- и Д2-рецепторов. Д2 намного меньше, но чувствительность их, установленная клиническими методами, к веществам их блокирующим (нейролептикам) в 100-1000 раз выше, чем у Д1 (хотя чувствительность Д1 к дофамину в 10 раз выше). Наиболее известный и первый нейролептик – аминазин (из химической группы фенотиазинов). Он снижает общий уровень тонуса ЦНС. Галоперидол – более специфический антагонист Д2-рецепторов. Блокирует маниакальные состояния и бред, не вызывая вялости и апатии. Плотность Д2 и Д4-рецепторов может возрастать, что повышает чувствительность постсинаптической мембраны к дофамину. Это характерно для шизофрении.
Среди стимуляторов выброса дофамина известна группа амфетаминов, к которым быстро развивается привыкание. Такой же неприятной особенностью характеризуется наркотик – кокаин. Он работает как блокатор обратного захвата дофамина, что истощает запасы медиатора.
СЕРОТОНИН
Серотонин близок в химическом отношении к катехоламинам. Образуется из незаменимой аминокислоты – триптофана, которого больше всего в молочных продуктах.
На периферии серотонин оказывает сосудосуживающее действие. Использующие его нейроны находятся в основном в ядрах шва (средняя линия продолговатого мозга, моста и среднего мозга). Большая часть аксонов идет к конечному, промежуточному и среднему мозгу.
По-видимому, естественный конкурент дофаминовой и норадреналиновой систем – снижает активность ЦНС. В сенсорных системах ограничивает распространение возбуждения по нейронным сетям, помогая концентрировать, фокусировать информацию. Аналогично эта система работает в ассоциативной коре. Возможно, серотонинергические нейроны включены в системы положительных эмоций и соответствующих навыков. Можно говорить о его седативном действии.
Основных рецепторов в ЦНС два: 5НТ1 и 5НТ2. Эти рецепторы могут связываться с галлюциногенами, например, ЛСД. Его действие нарушает восприятие, нормальную переработку информации. Эффект зависит от особенностей распределения рецепторов в мозгу человека и их чувствительности. ЛСД вызывает панические и параноидальные реакции, вероятность которых возрастает с увеличением дозы, на фоне эмоциональных переживаний.
При депрессии целесообразно усилить действие серотонина. Проще всего совместить это с усилением работы норадреналина и дофамина. Для этого блокируют фермент МАО. Более тонко можно устранять депрессии, сопровождаемые страхом, с помощью ослабления обратного захвата только серотонина (прозак или флуоксетин).
ГЛУТАМИНОВАЯ КИСЛОТА (ГЛУТАМАТ)
Глутаминовая кислота – главный возбуждающий медиатор в ЦНС. Представляет собой заменимую пищевую аминокислоту, которая, однако, плохо проникает через гематоэнцефалический барьер. Аминокислоты как медиаторы используются мозгом для быстрой передачи информации.
Глутаминовые рецепторы сложно устроены и для нормального функционирования нуждаются в небольшом количестве глицина. Глицин – аминокислота, медиатор, который обычно функционирует в тормозных синапсах. Его роль как соактиватора глутаминовых рецепторов осуществляется в очень низких концентрациях, в которых его собственные медиаторные свойства не проявляются.
Кроме того, включение рецепторов зависит от ионов Mg2+, которые до определенного уровня деполяризации блокируют работу рецептора. Включение рецептора означает не только вход в нейрон Na+, но и Ca2+, который оказывает на клетку мощное метаболическое воздействие. Поэтому при чрезмерной активации рецепторов может наблюдаться перевозбуждение, особенно в тех частях мозга, где гематоэнцефалический барьер ослаблен (гипоталамус, ромбовидная ямка продолговатого мозга).
Антагонисты глутаминовой кислоты оказывают тормозное действие и способны избирательно понижать патологическую активность мозга, например, при эпилепсии, болевых синдромах и т.д.
Некоторые антагонисты глутаминовых рецепторов (например, кетамин) являются мощными анальгетиками, но могут вызывать галлюцинации, затрагивающие осязательную сферу.
Инактивация глутаминовой кислоты осуществляется глиальными клетками, которые превращают ее в другие соединения, в том числе в ГАМК.
ГАМК – ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА
Это непищевая аминокислота. В ЦНС в основном участвует в работе тормозных синапсов. Рецепторы ГАМК обычно содержат Cl--каналы, но есть влияния и на К+-проницаемость.
В структуре рецептора есть места для присоединения других веществ, например, барбитуратов и бензодиазепинов, которые увеличивают время работы Cl--каналов и, следовательно, торможения.
Барбитураты применяют как средство наркоза и при тяжелых формах эпилепсии (фенобарбитал). Прием барбитуратов с алкоголем усиливает их действие, возможна передозировка и остановка дыхания. Сейчас чаще применяют бензодиазепины, в том числе как снотворные (нитразепам). При эпилепсии комбинируют бензодиазепины с антагонистами глутаминовой кислоты (ламотриджин). Это позволяет свести к минимуму побочные эффекты.
Как противосудорожные препараты применяют транквилизаторы (элениум, реланиум, тазепам, мезапам). Транквилизаторы не уменьшают психические расстройства (бред, галлюцинации, двигательные проявления), но уменьшают эмоциональную напряженность, тревоги, страхи.
Бензодиазепины применяют для облегчения абстинентного синдрома и ряда других проблем. Это связано с их механизмом действия: бензодиазепины связываются с активным центром, реагирующим на особые пептиды – эндозепины, которые блокируют ГАМК-рецепторы, т.е. не позволяют работать Cl--каналам, и вызывают возбуждение, страх. Таким образом, бензодиазепины – антагонисты эндозепинов.
ГАМК плохо проходит через гематоэнцефалический барьер, но, тем не менее, может использоваться при различных патологиях мозга, улучшая работу тормозных нейронов. Эффективны гибриды ГАМК с витаминами и производные ГАМК. Это группа ноотропов, к которой относится ноотропил (пирацетам). Эти препараты улучшают метаболические процессы в нейронах.
Еще один из тормозных медиаторов, активизирующий Cl--канал – глицин. Он эффективен при абстинентном синдроме, уменьшает возбуждение в стволе головного мозга, возможно, поэтому снижает риск инфаркта.
МЕДИАТОРЫ – ПЕПТИДЫ
Пептидные медиаторы – вещества, состоящие из небольшого количества аминокислот.
Наиболее изученной группой этих медиаторов являются опиоидные пептиды. Это аналоги природных соединений, обладающих анальгетическим, эйфорическим действием (морфиноподобные наркотики). Их делят на эндоморфины (4 аминокислоты), энкефалины (5 аминокислот), диморфины (8 аминокислот) и эндорфины (11 и более аминокислот). Выделено 3 типа рецепторов: мю (μ), дельта (δ) и каппа (κ). μ-рецептор взаимодействует в основном с эндоморфинами. Агонистом для них является морфий. δ взаимодействует в основном с энкефалинами, κ – с диморфинами. Первичная структура рецепторов совпадает на 50-70%.
Медиаторы образуются из общего белка-предшественника.
Основной механизм действия опиоидов в мозгу – пресинаптическое торможение выделения медиаторов. Механизм этот очень тонкий и сложный. Например, вещество Р снижает в спинномозговых нейронах активность Са2+-каналов, что уменьшает выброс глутамата, и передача болевых сигналов ослабляется.
В ядрах шва опиоиды блокируют ГАМК-ергические нейроны, которые тормозят серотониновые клетки. Серотониновые нейроны активизируются и ослабляют проведение боли в спинном мозгу.
Агонисты опиоидных рецепторов (морфин и другие соединения) вызывают мощное пресинаптическое торможение, поэтому могут обезболивать. Но этот эффект кратковременный. Нейроны, проводящие боль, увеличивают синтез аденилатциклазы, которая обеспечивает синтез цАМФ – посредника метаболизма нейрона. Поэтому для поддержания эффекта обезболивания приходится увеличивать количество морфия. Если морфий «убрать», количество цАМФ оказывается очень большим, и передача болевых и прочих сигналов резко возрастает – это абстинентный синдром.
Антагонистом опиоидных рецепторов является налоксон, который применяют при передозировке наркотиков.
Близко к пептидам-медиаторам по своим регуляторным свойствам примыкают нейрогормоны. Так, тиролиберин, влияющий на выделение гормонов щитовидной железы, одновременно активизирует эмоциональное поведение и бодрствование; холецистокинин (стимулятор желчного пузыря) может вызывать тревожность и страх; вазопрессин (регулятор деятельности почек) влияет на процессы запоминания; АКТГ (ключевой гормон стресса) стимулирует внимание, значит, участвует в механизмах памяти и улучшает метаболизм нейронов.
ЛИТЕРАТУРА:
1. Дубынин В.А., Каменский А.А., Сапин М.Р., Сивоглазов В.И. Регуляторные системы организма человека. М., Дрофа, 2003.
2. Костюк П.Г. Физиология центральной нервной системы. Киев, Высшая школа, 1979.
3. Ноздрачев А.Д., Баранников И.А. Общий курс физиологии человека и животных. Кн. 1. Физиология нервной, мышечной и сенсорной систем. М., Высшая школа, 1991.
4. Окс С. Основы нейрофизиологии. М., Мир, 1969.
5. Смирнов В.М. Физиология человека. М., 2001.
6. Физиология человека (ред. Покровский В.М., Коротько Г.Ф.), 2-е изд. М., Медицина, 2003.
7. Фомин Н.А. Физиология человека. М., ФиС, 1998.
8. Шмидт Р., Тевс Г. (Ред.) Физиология человека. Т. 1. Нервная система. М., Мир, 1996.
9. Шульговский В.В. Основы нейрофизиологии. М., Аспект-Пресс, 2000.
ВОПРОСЫ ДЛЯ САМОКОНТРОЛЯ:
1. Что такое рефлекс? Рефлекторная дуга? Рефлекторное кольцо?
2. Какие рефлексы Вы знаете?
3. Что такое нервный центр? Критерии определения нервных центров.
4. Какими свойствами обладает нервный центр?
5. Что такое координация нервных центров?
6. Механизмы координации.
7. Что такое доминанта? Какими свойствами она обладает?
8. Что такое обратные связи? На какие механизмы они опираются?
9. Что такое иерархия функций? Какие блоки участвуют в управлении движениями?
10. Как сокращаются мышцы?
11. Что такое двигательное качество? Какие механизмы лежат в основе?
12. Где записаны программы движений?
13. Чем отличаются вегетативные и соматические нервные центры?
14. Каковы особенности вегетативной регуляции?
15. Значение и особенности функционирования разных медиаторных систем.
16. Функции основных отделов ЦНС.