

БХ - 4 семестр / Разное / Ф
.pdfКафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
Общая
структура ДЦ Мх
16.11.2012 |
263 |
Коэффициент P/O
•P/O количественный показатель степени сопряжения
(эффективности) ОФ
•Коэффициент P/O отражает отношение кол-ва молекул Фн, пошедших на синтез АТФ к кол-ву поглощенных атомов О2
АДФ + Фн → АТФ
•При транспорте электронов с NADH в ДЦ образуется ≈3 молей АТФ
•При транспорте электронов с FADH2 в ДЦ образуется ≈2 молей АТФ
– |
NADH зависимые субстраты |
P/O = 3 |
– |
FADH2 зависимые субстраты |
P/O = 2 |
– |
Адреналин, аскорбат |
P/O = 1 |
16.11.2012 |
264 |
|
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
131 |

Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
Электрон-транспортная (дыхательная) цепь ЭТЦ (ДЦ)
16.11.2012 |
265 |
Функции ДЦ Мх
ДЦ Мх - общая финальная стадия БО всех аэробных клеток
1.За счет энергии окисления NADH+ H+ и FADH2 до
NAD+ и FAD в ДЦ образуется μН+,
2.Сопряжение ТД и ОФ - в сопрягающих комплексах (I, III
и IV) ДЦ Мх энергия транспорта электронов ( μН+) конвертируется в энергию АТФ и др виды:
3.Главный источник АТФ и др. видов биологически конвертируемой энергии аэробных клеток
Эн хим связи →Эн |
μН+ → др виды Эн |
(хим связи АТФ, механич, осмотич, световая, t ) |
|
16.11.2012 |
266 |
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
132 |

Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
Хемиосмотическое сопряжение
•Общий путь используется Мх, ХП и прокариотами для производства энергии на биологические нужды
солнце |
пища |
• Хемиосмотическое сопряжение
NADH, FADH2 отражает связь между
энергизованные электроны химическими реакциями («хеми»)
генерации АТФ и мембранным транспортом («осмотическое»)
|
|
μН+ |
|
|
|
|
|
|||
|
(Δ Ψ и |
рН+ ) |
|
• |
Процесс сопряжения протекает в |
|||||
|
|
|
|
|
|
|
|
две связанные стадии, каждая из |
||
Активный |
|
|
|
|
|
t |
которых происходит с участием |
|||
мембранный |
|
|
|
|
|
|||||
транспорт |
|
|
|
|
|
|
комплексов ДЦ внутренней |
|
||
|
|
|
|
|
|
мембраны Мх |
|
|||
|
|
|
|
|
|
|
|
|
||
|
|
|
|
Вращение |
|
|
|
|
|
|
|
|
|
жгутиков и |
|
Свечение |
|
|
|||
|
Синтез |
|
|
|
267 |
|||||
16.11.2012 |
|
|
|
|
|
|
|
|
||
|
АТФ |
|
др. |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
Структура протонного градиента
• μН+ состоит из Ψ и рН+
•
• Величина μН+ ≈ 180 мВ (≈ 250 кВ/см2
внутр мембраны Мх)
•рН+ составляет ≈ 1 рН
•Общая площадь внутр мембр Мх
клеток H.s. ≈ 100 000 м2
• Рассеить энергию μН+ (разобщить ОФ) можно
воздействуя на оба компонента
16.11.2012 268
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
133 |
Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
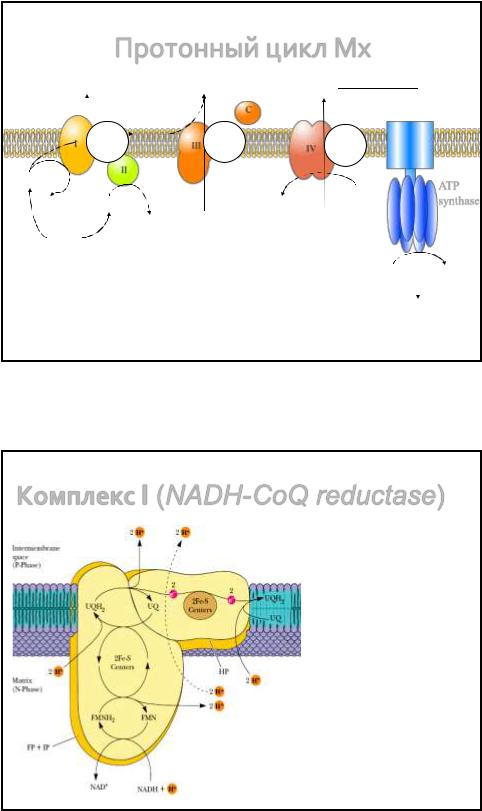
Протонный цикл Мх
4H+ 4H+ 2H+
|
|
|
F-цикл |
QH2 |
Q-цикл |
|
O-цикл |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
NADH + H+ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4H+ + O |
NAD+ |
|
FADH2 |
|
|
H O |
2 |
|
|
|
4H+ |
|
|
2 |
|
|
|
|
FAD |
2H+ |
|
2H+ |
||
|
TCA |
|
|
|
|
|
|
|
|
|
|||||
• В ДЦ имеется 3 цикла: F-цикл, Q-цикл и O-цикл. |
|||||||
• Протонные насосы формируют ΔμH+ |
|
ADP + Pi |
|||||
• Энергия ΔμH+ используется для совершения
полезной работы (синтез АТФ, осмотической, H+ механической и др.)
16.11.2012
ATP synthase
ATP
269
Комплекс I (NADH-CoQ reductase)
• Содержит: 1. FMN
2. FeS центры
(22-24 (Fe-S)
белка в 5-7 кластерах
• Акцептор электронов Ко Q
16.11.2012 |
270 |
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
134 |

Кафедра биохимии ГомГМУ, 2012 |
|
|
|
|
|
|
|
16.11.2012 |
|||
|
Коэнзим Q10 (КoQ10) или Убихинон |
||||||||||
|
|
|
|
O |
|
|
|
|
|
|
|
|
|
H3C |
O |
|
|
CH3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H3C |
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
CH3 |
10 |
|
|
|
• |
КoQ10 компонент расположенный в «середине» ДЦ |
|
|
||||||||
• транспортирует электроны с NADH, с сукцината, ацил-КоА и |
|||||||||||
|
α-глицерофосфата |
|
|
|
|
|
|
|
|
||
|
16.11.2012 |
|
|
|
|
|
|
|
|
271 |
|
|
|
|
|
|
КoQ10 |
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
|
|
H3C |
O |
CH3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H3C |
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
CH3 |
CH3 |
CH3 |
CH3 |
CH3 |
CH3 |
CH3 |
CH3 |
CH3 |
CH3 |
|
• |
Окисляется цитохромами |
|
|
|
|
|
||||
|
• |
Это коллектор электронов нескольких FAD- |
|
||||||||
|
|
зависимых ДГ |
|
|
|
|
|
|
|
||
|
16.11.2012 |
|
|
|
|
|
|
|
|
272 |
|
Введение в биохимию. Энзимология. |
|
|
|
|
|
|
|
||||
Биоэнергетика |
|
|
|
|
|
|
|
|
|
|
135 |

Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
Комплекс II (Сукцинат-КoQ редуктаза) или
СДГ
• Комплекс II содержит
FAD и 7-8 Fe-S белков в
3 кластерах
• Комплекс II СДГ,
одновременно фермент ЦТК и интегральный белок внутренней мембраны Мх
• Масса СДГ от 100 до 140 kD
16.11.2012 |
273 |
Комплекс II и III
•КoQ акцептирует электроны с обеих комплексов I и II и переносит на комплекс III.
16.11.2012 |
274 |
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
136 |

Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
Комплекс IV: Цитохром c Оксидаза
• Комплекс IV назван
Цитохром c Оксидазой потому
что акцептирует
электроны с цитохрома с и направляет для 4-х электронного восстановления O2 до H2O
16.11.2012 |
275 |
Комплекс V H+-АТФ-аза
(АТФ-синтаза )
• |
Ионные градиенты на |
|
внутренней мембране - форма |
|
хранения биологически |
|
конвертируемой энергии, |
|
которая совершает полезную |
|
работу, когда поток ионов идет |
|
в обратном направлении |
• Обратный поток H+ против его |
|
|
электрохимического градиента |
|
через АТФ-синтазу – источник |
|
синтеза АТФ из АДФ и Фн |
• |
Этот вездесущий фермент |
|
играет роль турбины, |
|
превращающей энергию |
|
протонного градиента в АТФ |
16.11.2012 |
276 |
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
137 |

Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
ATФ/AДФ - транслоказа
• Транспорт АТФ из Мх (ч/з ATФ/AДФ транслоказу) происходит в мембране Мх по электрохимическому
16.11.2012 потенциалу (∆µH+) 277
Функционирование ДЦ
16.11.2012 |
278 |
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
138 |
Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
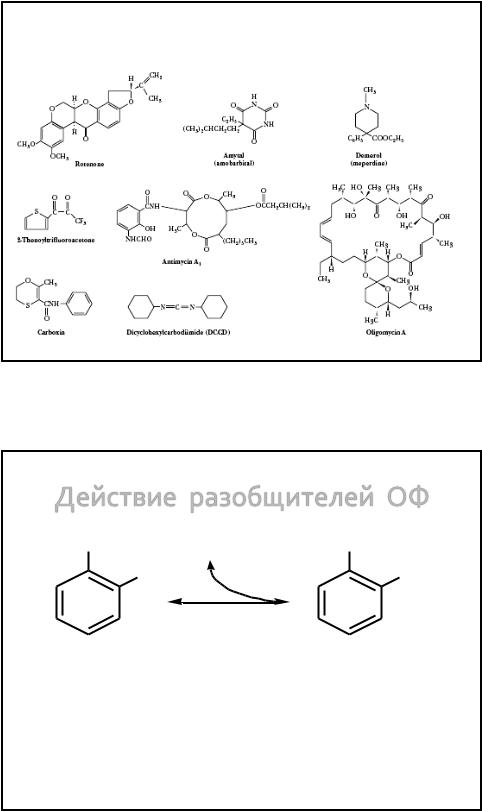
The Structures of Several Inhibitors of ETC
and OP
16.11.2012 |
279 |
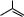
Действие разобщителей ОФ
O- |
H+ |
OH |
|
NO2 |
NO2 |
|
|
|
|
NO2 |
NO2 |
||
•2,4-динитрофенол (2,4-ДНФ), как слабая кислота
связывает протон в межмембранном пространстве и
переносит его в матрикс Мх (рассеивая энергию
ΔμH+ ) |
280 |
16.11.2012 |
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
139 |

Кафедра биохимии ГомГМУ, 2012 |
16.11.2012 |
2,4-ДНФ снижает ΔμH+
H+ |
H+ |
H+ |
+ |
|
+ |
H+ |
H+ |
+ |
|
+ |
+ |
+ |
|
+ |
|
Межмембранно |
H+ |
|
H+ H H+ |
H |
+ |
H+ |
H H+ |
H |
+ |
H H+ |
H |
25:1 H |
|
||
е пространство |
H+ H+ |
H+ |
H+ |
H |
|
|
H+ |
H |
H+ |
|
|
||||
|
|
|
|
|
|
- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
 NO2
NO2
NO2
NO2
O2N
OH
Матрикс Мх
|
H+ |
16.11.2012 |
281 |
Биоэнергетика Дж. Николс. М. Мир. 1985
16.11.2012 |
282 |
Введение в биохимию. Энзимология. |
|
Биоэнергетика |
140 |