

Особенности морфологического строения печени позволяют ей эффективно удалять и фиксировать ионы аммония.
П осле
приема пищи концентрация ионов аммония
в портальной вене - 0.3 мM. Когда кровь
проходит по сосудам печени, концентрация
ионов аммония в ней может повышаться
до1 мM. Однако, когда кровь покидает
печень и входит в системную циркуляцию,
концентрация понижается до 0.02 мM. Эти
возможности печени по усвоению аммиака
связаны с особенностями морфологического
строения печени
осле
приема пищи концентрация ионов аммония
в портальной вене - 0.3 мM. Когда кровь
проходит по сосудам печени, концентрация
ионов аммония в ней может повышаться
до1 мM. Однако, когда кровь покидает
печень и входит в системную циркуляцию,
концентрация понижается до 0.02 мM. Эти
возможности печени по усвоению аммиака
связаны с особенностями морфологического
строения печени
Гепатоциты расположенные вокруг капилляров портальной системы после приема пищи получают кровь, богатую питательными веществами. Эти клетки обогащены глутаминазой и ферментами синтеза мочевины. Глутаминаза аллостерически активируется ионами аммония, так что ионы аммония, поступающие в гепатоциты, активируют фермент, который превращает глутамин в ионы аммония и глутамат. Основная масса ионов аммония удаляется из циркуляции перипортальными гепатоцитами. Кm карбомоилфосфатсинтетазы для аммиака примерно 1 мM. Карбомоилфосфатсинтетаза I и глутаминаза аллостерически активируются N-ацетилглутаматом.
Перивенозные гепатоциты образуют один из слоев клеток, что окружают центральную вену. Эти клетки обогащены глутаминсинтетазой, с низким значением Кm для ионов аммония. Поэтому ионы аммония, которые не превратились в мочевину в перипортальных клетках, накапливаются в составе глутамина в перивенозных клетках.
Орнитиновый цикл мочевинообразования - главный механизм обезвреживания аммиака.
Печень единственный
орган, клетки которого содержат все
ферменты синтеза мочевины и, следовательно,
являются главным местом синтеза мочевины
в организме. Ферменты, участвующие в ее
синтезе расположены в митохондриях и
цитозоле.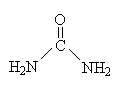 Синтез мочевины – пример циклического
метаболического пути. Орнитин представляет
собой пример непротеиногенной
диаминомонокарбоновой кислоты, которая
играет роль своеобразного катализатора
процесса, начиная этот процесс в качестве
акцептора карбамоилфосфата и, в
заключительной реакции, после удаления
молекулы мочевины она вновь регенерирует.
Расходуются в процессе мочевинообразования
аспарагиновая кислота и ион аммония (
источники аминогрупп молекулы мочевины),
диоксид углерода (из бикарбоната) и АТФ.
Синтез 1 моля мочевины требует 3 молей
АТФ.
Синтез мочевины – пример циклического
метаболического пути. Орнитин представляет
собой пример непротеиногенной
диаминомонокарбоновой кислоты, которая
играет роль своеобразного катализатора
процесса, начиная этот процесс в качестве
акцептора карбамоилфосфата и, в
заключительной реакции, после удаления
молекулы мочевины она вновь регенерирует.
Расходуются в процессе мочевинообразования
аспарагиновая кислота и ион аммония (
источники аминогрупп молекулы мочевины),
диоксид углерода (из бикарбоната) и АТФ.
Синтез 1 моля мочевины требует 3 молей
АТФ.
Синтез карбамоилфосфата – ключевая реакция синтеза мочевины
Карбамоилфосфат синтезируется в митохондриях в комплексной реакции, которая катализируется карбамоилфосфат синтетазой I. Этот аллостерический фермент активируется N –ацетилглутаминовой кислотой. Потребление 2 молекул АТФ делает синтез карбамоил-фосфата необратимой реакцией. Часть этой энергии депонируется в виде энергии макроэргической связи (смешанная ангидридная связь).
Карбамоилфосфат синтетаза I ( КФС-1) катализирует образование карбомоилфосата в митхондриях. Из цитозоля выделена карбомоилфосфатсинтетаза-2 (КФС-2), которая катализирует образование карбомоилфосфата, используемого для синтезе пиримидиновых нуклеотидов.
Образование цитруллина – первая реакция орнитинового цикла.
Цитруллин подобно орнитину - основная аминокислота, которая не кодируется генетически.

Орнитиновый цикл синтеза мочевины
Образование цитруллина происходит благодаря использованию высокоэнергетического фосфата карбамоилфосфата и катализируется орнитин-карбамоил трансферазой (ОКТ). Продукт реакции цитруллин транспортируется в цитозоль, где завершается образование мочевины.