

Биологическое окисление 1
Лектор
д.м.н., проф. Грицук А. И.
зав. кафедрой биохимии Гомельского государственного медицинского университета
ЛФ
07/06/19

Содержание
Биоэнергетика: История развития
Биологическое окисление
ЦТК (Цикл Кребса)
07/06/19

Биоэнергетика
Термин введен лауреатом Нобелевской премии Альбертом Сент-Дьерди
Биоэнергетика - раздел биохимии изучающей механизмы и пути превращения энергии в живых системах
07/06/19

История учения о БО
Античные авторы: Учение о 4 стихиях обсуждали роль воздуха в БО Платон (воздух необходим для поддержания
внутреннего огня) «Могущественные боги, предоставив нам, немощным пищу, разделили тело наше каналами, чтобы оно могло орошаться как бы из некого идущего сверху потока… Получив орошение и освежение, оно имеет возможность питаться и жить. Ибо когда воздух входит внутрь и выходит вон, то и соединенный с ним внутренний огонь охватывает пищу и питье, расплавляя их, разлагает на мелкие частицы и затем приносит к жилам» Платон
Аристотель (воздух для охлаждения внутренностей и крови)
XVII в. Georg Ernst Stahl создал теорию флогистона.
1770 годы . Carl Scheele и Joseph Priestley открыли
бесфлогистонный воздух, который впоследствии А. Лавуазье назвал кислородом
07/06/19

Antoine Lavoisier
В конце XVIII в. A. Лавуазье ввел в
химические исследования количественный метод и создал кислородную теорию горения
A. Лавуазье установил сходство горения и дыхания на основании количественного анализа конечных продуктов
07/06/19

Теория активации кислорода
|
В 1840 Ф. Шёнбайн открыл озон, более активную форму О2 |
||||||||||||
|
В конце XIX в. – почти одновременно A.Н. Бах (Россия) и |
||||||||||||
|
К. Энглер (Германия) создали теорию активации кислорода : |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
O |
O |
|
||||||
|
A + |
|
|
|
|
|
|
A |
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
O |
|
O |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
Гипотетическая |
|
|
|
|
|
|
|
|
|
|
|
|
|
форма озона |
|
|
|
|
O |
|
|
|
|
|
|
|||||
|
A |
|
|
|
+ SH2 |
|
|
S + A + H2O2 |
|
||||
|
|
|
|
|
|
|
|||||||
|
|
O |
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|

Критическая оценка теории Баха-Энглера
1.Не была найдена высокая активность оксигеназ в живых организмах.
2.Не была найдена высокая концентрация H2O2 в живых организмах.
3.Были обнаружены ферменты деградации H2O2 в живых организмах (каталаза и пероксидаза).
H2O2 H2O + O |
(каталаза) |
H2O2 + 2GSH 2H2O + G-S-S-G |
(GSH пероксидаза) |
07/06/19 |
|

|
|
Теория Палладина-Виланда |
||||||
|
1903 – Н. Бор, создал теорию строения атома (ядро, электроны). |
|||||||
|
|
Возникло иное представление об О-В процессах |
||||||
|
|
(бескислородное окисление) |
|
|||||
|
|
1912 –В.И. Палладин и Г. Виланд создали теорию «активации |
||||||
|
|
водорода», предполагающую наличие 2-х стадий: |
||||||
|
|
Анаэробная стадия: SH2 + R = S + RH2 (см рис.) |
||||||
|
|
Аэробная стадия : RH2 + 1/2 O2 R + H2O. |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
O |
OH |
||||
|
SH2 + |
|
|
|
|
|
S + |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|||
|
|
|
|
|
|
|||
|
|
|
|
|
|
OH |
||
|
|
|
|
|
|
|
||
|
|
|
O |
|||||
|
|
quinone |
hydroquinone |
|||||

Хромогены и гистогематины
Переносчики электронов были названы хромогенами из-за окраски зависящей от О-В состояния
В качестве хромогенов позднее идентифицированы коферменты FMN, FAD, NAD+, NADP+.
1925 –открыты гистогематины (цитохромы)
1932 – акад В.А. Энгельгардт открыл сопряженное окислительное фосфорилирование
(ADP + Pi ATP).
07/06/19
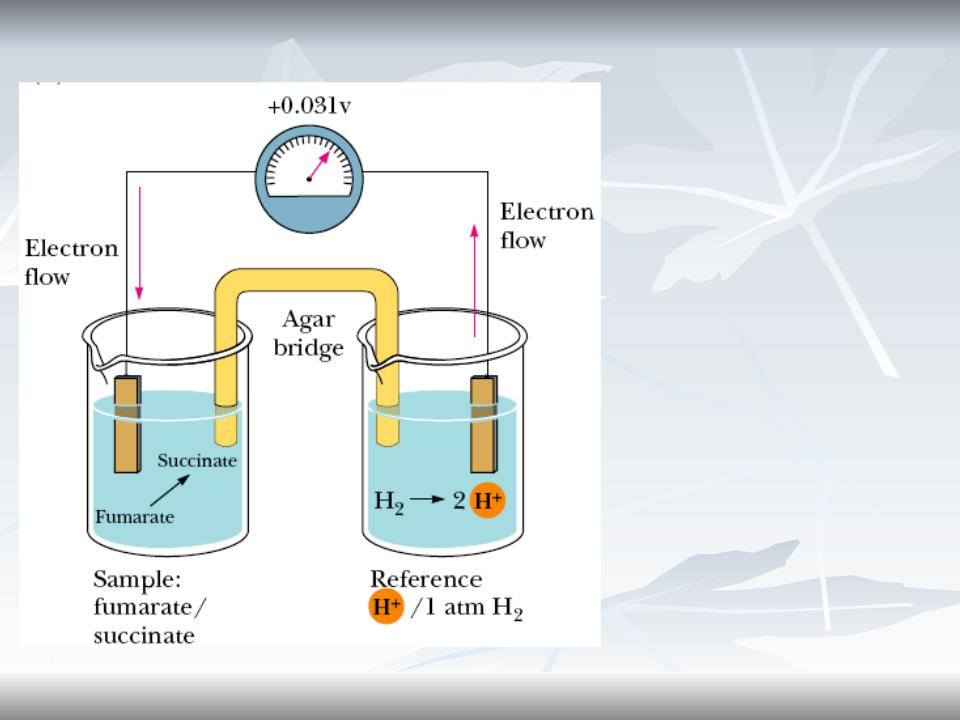
Редокс-реакции, редокс потенциал
Реакции переноса электронов от донора к акцептору.
Редокс потенциал создает напряжение в цепи.
Редокс потенциал пары H+:H2 принят за 0.
07/06/19
ATP
NH2
N
 N
N
O |
O |
O |
|
|
N |
N |
|
|
|
||||
-O P O |
P |
O P |
O |
|
O |
|
|
|
|
|
|
|
|
O- |
O- |
O- |
|
H |
H |
|
|
|
|
H |
OH |
H |
|
|
|
|
|
OH |
|
ATP – основной макроэрг всех живых клеток.
Содержит 2 макроэргические связи.
07/06/19