

ДЕРЖАВНИЙ ВИЩИЙ НАВЧАЛЬНИЙ ЗАКЛАД
«ЗАПОРІЗЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ»
МІНІСТЕРСТВА ОСВІТИ І НАУКИ УКРАЇНИ
О. К. Фролов, В. В. Копійка, Є. Р. Федотов, Р. О. Литвиненко
ЦИТОЛОГІЯ
Методичні вказівки до лабораторних занять
для студентів освітньо-кваліфікаційного рівня «бакалавр» напряму підготовки «Біологія»
Затверджено
Вченою радою ЗНУ
Протокол № від
Запоріжжя
2014
УДК 576. 3 (076)
ББК 28.05я73
Ц 747
Цитологія: методичні вказівки до лабораторних занять для студентів освітньо-кваліфікаційного рівня «бакалавр» напряму підготовки «Біологія» / Уклад.: О. К. Фролов, В. В. Копійка, Є. Р. Федотов, Р. О. Литвиненко. – 3-тє вид., розшир. та доповн. – Запоріжжя: ЗНУ, 2014. – 51 с.
Методичні вказівки складені відповідно до навчальної та робочої програм із дисципліни «Загальна цитологія». Вони призначені для студентів денної та заочної форм навчання біологічного факультету. Основна їх мета полягає в ознайомленні студентів із основними ознаками клітинного рівня організації життя, яка досягається шляхом аналізу структури і функції біомолекул, органоїдів, причому розгляд клітинних структур іде в філогенетичному аспекті. Структура методичних вказівок побудована за оптимальною педагогічною схемою: цільові установки, перелік питань для самопідготовки, навчальні завдання, питання для самоконтролю за успішністю засвоєння знань на заняттях. У ході виконання практичних завдань студенти повинні зробити протокольні записи у формі рисунків, схем, аналітичних висновків, які зараховуються викладачем у кінці заняття.
Рецензент В. Д. Бовт
Відповідальний за випуск О. К. Фролов
ЗМІСТ
Тема 1. Мікроскоп, приготування препаратів і техніка мікроскопування….........4
Тема 2. Методи дослідження у цитології..................................................................6
Тема 3. Еволюція клітинних форм життя..................................................................9
Тема 4. Біологічні мембрани ……………………………………………….…......12
Тема 5. Міжклітинні контакти. Спеціалізовані структури плазматичної мембрани. Клітинні оболонки………………………………………………….....14
Тема 6. Ядро клітини. ДНК та білки хроматину. Рівні організації хромосом………………………………………………………………………........16
Тема 7. Ядро клітини. Метафазні хромосоми. Статевий хроматин. Каріотип…………………………………………………………………………......19
Тема 8. Ядро клітини. Негенетичні структури. Оболонка, каріоплазма, матрикс. Ядерце та рибосоми………………………………………………….......................22
Тема 9. Органели цитоплазми. Гіалоплазма. Вакуолярна система…………......25
Тема 10. Органели цитоплазми. Мітохондрії. Пероксисоми……………….......27
Тема 11. Опорно-рухова система клітини………………………….………..........31
Тема 12. Розмноження еукаріотичних клітин. Мітоз………………………….....36
Тема 13. Відхилення від звичайного протікання мітозу: ендомітоз, ендоредуплікація, багатоядерність. Відкритий та закритий мітози. Багатополюсність мітозу та порушення окремих ниток веретена.………….......41
Тема 14. Розмноження організмів……………………………………………........43
Тема 15. Мейоз: цитологічні та генетичні закономірності…………………........44
Рекомендована література……………………………………………………..…..49
темА 1
Мікроскоп, ПРИготування препаратів і техніка мікроскопУвання
МЕТА ЗАНЯТТЯ: Ознайомити студентів із інструментами, реактивами, що використовуються на лабораторних заняттях; вивчити будову оптичного мікроскопа і правила користування ним; вивчити техніку приготування постійних і тимчасових препаратів.
Питання для самопідготовкИ:
1. Історія створення та удосконалення оптичного й електронного мікроскопів.
2. Будова світлового мікроскопа.
3. Правила роботи зі світловим мікроскопом.
4. Підготовка препаратів до мікроскопування.
Навчальні завдання
ЗАВДАННЯ 1. Правила користування світловим мікроскопом.
1. Установка мікроскопа: мікроскоп поставити колонкою до себе і центрувати об’єктив малого збільшення (повертати револьвер до клацання).
2. Освітлення поля зору: діафрагма відкрита повністю; дивлячись в окуляр повертати дзеркало вбік джерела світла до максимально яскравого і рівномірного освітлення поля зору. Після цього рухати мікроскоп не можна. При автономному освітленні мікроскопа — освітлення встановлюють згідно правила Келера: перевести освітлення на мінімум, закрити діафрагму і відцентрувати отвір діафрагми в центр поля зору шляхом регулювання гвинтами конденсора, після чого повністю відкрити діафрагму і поставити спіраль лампи в центр поля зору поворотами її цоколя. Після чого регулювати освітлення відповідно збільшення мікроскопа і цитологічної потреби мікропрепарату.
3. Розташування препарату на столику: покласти препарат на предметний столик мікроскопа покривним склом догори. Досліджуваний об’єкт повинен знаходитися точно під об’єктивом малого збільшення.
4. Вивчення об’єкта при малому збільшенні: дивлячись збоку на препарат, опустити тубус за допомогою макрогвинта так, щоб відстань між фронтальною лінзою об’єктива і покривним склом препарату була близько 0,5 см. Потім, дивлячись в окуляр, за допомогою макрогвинта підняти тубус до появи чіткого зображення.
5. Вивчення об’єкта при великому збільшенні: для переведення мікроскопа на велике збільшення об’єкт встановити в центр поля зору на малому збільшенні. Підняти тубус, повернувши макрогвинт до себе на півоберта. Поворотом револьверної системи до клацання встановити об’єктив великого збільшення. Після цього, дивлячись збоку на препарат, опустити тубус за допомогою макрогвинта так, щоб відстань між фронтальною лінзою і препаратом була менше 1 мм. Дивлячись в окуляр, повільно піднімати тубус до появи зображення. Для точного фокусування і вивчення препарату гвинт необхідно повертати в глибину не більше, ніж на півоберта.
6. Отримання зображення під імерсією: перед отриманням зображення під імерсією необхідно відцентрувати препарат на малому і великому збільшеннях, додати краплю імерсійної олії на покривне скло препарату та відцентрувати імерсійний об’єктив. Дивлячись збоку, за допомогою макрогвинта обережно опустити імерсійний об’єктив у краплю імерсійної олії, потім, дивлячись в окуляр, за допомогою макрогвинта обережно трохи підняти об’єктив, щоб переконатися, що фокус не пройдений, а потім також дуже обережно опускати імерсійний об’єктив до появи чіткого зображення. Остаточне встановлення фокуса зображення і вивчення препарату в глибину здійснювати за допомогою мікрогвинта.
Після роботи з імерсійною системою обов’язково видалити імерсійну олію з об’єктива мікроскопа та препарату!
7. Переведення мікроскопа на мале збільшення: при переведенні мікроскопа з великого збільшення на мале, не підіймаючи тубуса, повернути револьверну систему до установки об’єктива малого збільшення.
8. Завершення роботи: закінчивши роботу, мікроскоп перевести на мале збільшення, зняти препарат.
ЗАВДАННЯ 2. Принцип приготування постійних препаратів для світлового мікроскопування.
Складіть схему приготування цитологічних і гістологічних препаратів. У схему включити такі етапи: 1) взяття зразка; 2) фіксацію; 3) відмивання від фіксатора і зневоднення в батареї спиртів зростаючої концентрації і ксилолі; 4) замкнення у парафін і приготування блоків; 5) нарізку препаратів на мікротомі; 6) депарафінізацію в ксилолі і спиртах понижуючої концентрації; 7) фарбування водорозчинними барвниками; 8) відмивання барвника і зневоднення в спиртах зростаючої концентрації; 9) проведення через ксилол і замкнення у бальзам між предметним і покривним склом. Дайте відповідне пояснення кожному етапу.
ЗАВДАННЯ 3. Волокна вати і пухирці повітря під мікроскопом.
Приготуйте тимчасовий препарат, що складається з волокон вати і води з пухирцями повітря. Знайдіть на малому збільшенні (об. 10х; ок. 10х) і на великому (об. 40х; ок. 10х) пухирець повітря з волокнами вати, вивчить їх в об’ємі, замалюйте в альбом на великому збільшенні. Дайте відповідне пояснення мети такого дослідження.
ЗАВДАННЯ 4. Клітини крові жаби.
Мазок крові жаби. Фіксація метиловим спиртом, фарбування азур-еозином за Романовським-Гімза.
Знайдіть на малому збільшенні клітини крові жаби. Роздивіться їх морфологію під імерсією. Більшість клітин препарату – еритроцити. У амфібій вони мають овальну форму й овальне щільне ядро, забарвлене азуром у синьо-фіолетовий колір. Цитоплазма цих клітин фарбується еозином в оранжево-червоний колір за рахунок гемоглобіну. Крім еритроцитів знайдіть лейкоцити. Лімфоцити мають округлу форму, розмір менше за еритроцит, із щільним округлим ядром і вузьким обідком блакитної (базофільної) цитоплазми. Рідко зустрічаються еозинофіли. Це округлі клітини, за розміром менші, ніж еритроцити, із 3-4-сегментним щільним ядром і яскраво-жовтогарячою зернистістю в цитоплазмі. Між еритроцитами зустрічаються найменш дрібні клітини – тромбоцити. Вони мають округлу форму, округле ядро і блідо-фіолетову цитоплазму. Замалюйте по одній клітині: еритроцит, лімфоцит, тромбоцит, еозинофіл (у випадку знаходження). У висновках порівняйте з еритроцитами ссавців, дайте біологічне значення ароморфозів і ідіоадаптацій структурно-функціональних відмінностей.
Питання для самоКОНТРОЛЮ:
1. Роздільна здатність світлового та електронного мікроскопів.
2. Характеристика загального зображення світлового мікроскопа.
3. Як вивчити об’єкт дослідження при переході з малого збільшення на велике?
4. Який ефект імерсійної олії?
5. Артефакти постійних та тимчасових препаратів і засоби їх усунення.
6. Вимоги до фіксуючих розчинів, механізм їх дії.
7. Приготовані постійні препарати поганої прозорості, мутні. Які імовірні причини?
ТЕМА 2
МЕТОДИ ДОСЛІДЖЕННЯ У ЦИТОЛОГІЇ
МЕТА ЗАНЯТТЯ: вивчити методи дослідження у цитології, засвоїти принципи будови світлового, електронного та люмінесцентного мікроскопів.
Питання для самопідготовкИ:
1. Макросистема живих організмів, у тому числі неклітинних форм життя.
2. Стислий нарис історії цитології.
3. Сучасні положення клітинної теорії.
4. Загальний план будови клітин про- та еукаріот.
5. Нові методи у світловій мікроскопії (флуоресцентний, радіоімунний, імуноферментний та ін.) та електронної мікроскопії (заморожування і сколювання та ін.).
6. Вивчення будови і функцій клітин комплексом методів: електронної мікроскопії, біохімії, ультрацентрифугуванням.
Навчальні завдання
ЗАВДАННЯ 1. Методи дослідження в цитології.
Скласти загальну схему ходу проміння у світловому, електронному (просвітлюючий, скануючий), люмінесцентному мікроскопах.
ЗАВДАННЯ 2. Морфометрія як метод кількісного дослідження мікроструктур.
Ознайомтеся з одним із методів кількісної оцінки мікроструктур, що є необхідною умовою отримання об’єктивних даних про їх стан у нормі, при експериментальному впливі та у разі патології. Морфометрія включає сукупність прийомів та методів визначення геометричних характеристик досліджуваних об’єктів. Об’єктами можуть бути зображення препаратів (зрізи, мазки, відбитки), а також мікрофотографії. Вимірювання кількості структур, їх площі, діаметра, периметра та інших показників проводиться за допомогою спеціальних сіток, методу обчислення вагових співвідношень шляхом зважування. Вимірювання структур у світлових мікроскопах виконують за допомогою окуляр-мікрометру, вставленого в окуляр мікроскопа.
Для використання морфометричних методів не потрібно спеціального оснащення. Одним із варіантів планіметричного методу є «клітинний» або «сітковий» метод, коли в окуляр вставляється окуляр-мікрометр у вигляді лінійки або сітки Автандилова. Насамперед встановлюють, чому відповідає одна поділка окуляр-мікрометра при відповідному збільшенні мікроскопа, наприклад, об. 100х; ок. 7х. Для цього на предметний столик встановлюють об’єкт-мікрометр, виставляють його лінійні розміри в центр поля зору при даному збільшенні і сполучають лінійні розміри об’єкт-мікрометра (початок лінійки) із лінійними розмірами окуляр-мікрометра (край сітки Автандилова). Під мікроскопом визначають число поділок (а) об’єкт-мікрометра, що відповідають кількості поділок окуляр-мікрометра (в). Після цього знаходять ціну поділок використаних вставок за формулою: р = а × с / в, де с – ціна поділки об’єкт-мікрометра. Наприклад, об’єкт-мікрометр має ціну поділки 0,01 мм або 10 мікрометрів (мкм). 5 поділок об’єкт-мікрометра (5х10=50 мкм) співпадає з 8 квадратами сітки. Отже, ціна поділки сторони квадрата сітки Автандилова буде складати 50:8= 6,25 мкм.
Діаметр досліджуваної клітини визначають накладанням на мікропрепарат (на клітину, що вимірюється) квадратної сітки або лінійки з відомою ціною поділки. Потім підраховують кількість повних і неповних квадратів, які відповідають діаметру досліджуваної структури і, які будуть являти собою діаметр клітини в умовних одиницях (у.о.). Визначають два діаметри клітини (в умовних одиницях) – максимальний і мінімальний, після чого встановлюють його середнє значення, яке переводять у мікрометри, перемножуючи отримане значення в умовних одиницях на ціну поділки сторони квадрата сітки Автандилова. Наприклад, максимальний діаметр клітини складає 1,3 у.о., мінімальний – 1,1 у.о., середній діаметр – 1,2 у.о. або 1,2 х 6,25 мкм = 7,5 мкм.
За допомогою морфометричної сітки зробіть заміри розмірів 100 лімфоцитів у мазку крові. З розміром лімфоцитів пов’язана їх функціональна активність.
Складають варіаційний ряд розмірних класів лімфоцитів із інтервалом 1 мкм. Їх об’єднують у групи: малі, з діаметром клітини до 6,5 мкм (КЛ≤6,0 мкм); середні – 7-9 мкм (КЛ 7-9 мкм); великі – 10 і більше мкм (КЛ≥10 мкм). За даними О. К. Фролова та співавторів (2004) лімфоцити з діаметром до 6,5 мкм та частина КЛ≥10 мкм є активованими в організмі на момент взяття крові. Стан імунної системи визначають за частотою КЛ≤6,0 мкм, які складають ранні постпроліферативні зрілі активовані лімфоцити з високою міграційною активністю, а також за обліком зрушень лімфоцитарної формули крові вліво (вихід у рециркуляцію незрілих імунобластів) – за частотою великих розмірних класів лімфоцитів (КЛ≥10 мкм).
ЗАВДАННЯ 3. Основні структурно-функціональні розходження про- і еукаріот.
-
Заповніть таблицю принципових розходжень у загальному плані будови про- та еукаріот.
|
№ |
Ознаки |
Прокаріоти |
Еукаріоти |
|
1 |
Розмір клітин, мкм |
|
|
|
2 |
Кількість клітин |
|
|
|
3 |
Типи рибосом |
|
|
|
4 |
Інші органели |
|
|
|
5 |
Клітинні оболонки |
|
|
|
6 |
Тип будови джгутиків |
|
|
|
7 |
Фотосинтез |
|
|
|
8 |
Фіксація азоту |
|
|
|
9 |
Організація геному |
|
|
|
10 |
Тип живлення |
|
|
Питання для самоКОНТРОЛЮ:
1. Охарактеризуйте основні історичні етапи розвитку мікроскопічної техніки.
2. Назвіть основні методологічні підходи до вивчення клітини.
3. Роздільна здатність світлової мікроскопії (максимальне збільшення та засоби його підвищення).
4. Гістохімічні барвники, які використовуються в цитології.
5. Застосування люмінесцентних барвників у цитології. Переваги перед гістохімічними та радіоімунними методами.
6. Застосування проточних цитометрів у цитологічних дослідженнях.
7. Біотехнологічні методи в цитології (моноклональні антитіла та ПЛР-діагностика).
8. Основні принципи препаративної цитології (руйнування клітин, ультрацентрифугування та вивчення окремих фракцій).
9. Які положення клітинної теорії Т. Шванна, Р. Вірхова підтверджені положеннями сучасної цитології? Які залишені?
10. Вкажіть загальні риси (ознаки) структурно-функціональної будови клітин про- та еукаріот.
ТЕМА 3
ЕВОЛЮЦІЯ КЛІТИННИХ ФОРМ ЖИТТЯ
МЕТА ЗАНЯТТЯ: Пізнати основні структурно-функціональні механізми різноманіття та еволюції клітинних форм життя основного клітинного вчення.
Питання для самопідготовкИ:
1. Ендосимбіотичні теорії еволюції еукаріотичних клітин.
1.1. Виникнення тригеномної клітини – попередника царства грибів і тварин.
1.2. Виникнення квадригеномної клітини – попередника царства рослин.
2. Біологічні (структурно-функціональні) можливості до прогресивної еволюції про- та еукаріот.
3. Зальний план будови усередненої еукаріотичної клітини.
Навчальні завдання
ЗАВДАННЯ 1. Філогенетичні етапи еволюції клітин еукаріот (відповідно до ендосимбіотичної гіпотези).
Ознайомтесь із гіпотетичною схемою, що включає: прокаріотичну екологічну систему; тригеномну еукаріотичну клітину; квадригеномну еукаріотичну клітину.
При упорядкуванні схеми використовуйте такий гіпотетичний (проте який має вагомі докази) ланцюг еволюційних подій. У середині Архею метаболічне різноманіття досягло великої досконалості у всіх областях біосинтезу, яке могло б виникнути в елементарній клітинній системі: клітини могли синтезувати або утилізувати майже всі складні органічні речовини. Наслідком цього метаболічного різноманіття було виникнення екосистем: продуценти, консументи декількох порядків, редуценти. Проте прокаріотична екосистема виявилася безвихідною гілкою еволюції, тому що кожна прокаріотична клітина була вже метаболічно вузькоспеціалізованою. Еволюційний стрибок був пов’язаний із серією послідовних ендосимбіозів із утворенням багатофункціональної еукаріотичної системи.
Тригеномна еукаріотична клітина – прообраз клітин тварин і грибів – виникла з 3-х ендосимбіонтів. Нуклеоцитоплазма утворилася з великих анаеробних мікроорганізмів – хазяїв, але спроможних витримати високі температури і кислотність завдяки тому, що їх ДНК була захищена білками-гістонами. З ними ввійшли в ендосимбіоз (проникли усередину) аеробні мікроорганізми, згодом перетворившись у мітохондрії, що призвело до ефективності енергетичного обміну і синтезу нових складних органічних речовин, зокрема стероїдів, із якими пов’язане удосконалення багатьох метаболічних процесів: синтез гормонів, ліпоїдів, спроможність до піно- та фагоцитозу.
Анаеробна
бактерія Thermaplasma
acidophylia
нуклеоплазма амебоїд тригеномна
еукаріотична клітина тварини
гриби





1/
нуклеоцитоплазма 2/
мітохондрії 3/
ЦОМТи
Енодосим-біоз Енодосим-біоз
аеробна
бактерія (проміто- хондрія)
Спіро-плазма



Найост
Наступним еволюційним кроком до еукаріотичної клітини було набуття внутрішньо- і позаклітинної рухливості: центрів організаторів мікротрубочок. Їх привнесли прокаріотичні мікроорганізми за типом спіроплазм, що мають внутрішньо трубочкову систему.
Найостанніша еволюційна подія в серії послідовних ендосимбіозів – набуття еукаріотами спроможності до фотосинтезу. Для цього гетеротрофні еукаріоти фагоцитували фотосинтезуючих прокаріот, але не перетравлювали їх остаточно. Останні надалі еволюціонували в пластиди.
тригеномні гетеротрофні еукаріотичні клітини
квадригеномні аутотрофні еукаріотичні клітини
рослини Ендосим- біоз



фотосинтезуючі
прокаріоти ціанобактерії хлоробактерії
Вказати анаболічні та катаболічні еволюційні надбання кожного етапу ендосимбіозу.
ЗАВДАННЯ 2. Філогенетичні напрямки еволюції еукаріотичних клітин царства тварин.
Від предкової джгутикової клітини найпростіших складіть схему еволюції найпростіших у фізичних межах однієї клітини за рахунок диференціювання внутрішньоклітинних структур, приведіть до появи багатоклітинних тварин.
При упорядкуванні схеми використовуйте такі наукові дані.
Предкова джгутикова клітина Найпростіших
Колоніальна
багатоклітинність
Диференціювання
клітин




































Багатоклітинні
тварини










Колоніальні
джгутикові
поліплоїдія
та диференціювання
внутрішньоклітинних структур
інфузорії джгутикові споровики саркодові

«Глухий кут» еволюції еукаріот
Поліфункціональність одноклітинних еукаріотичних організмів стала основою їх подальшої еволюції. Проте і еукаріотам на початку не вдалося уникнути безвихідних еволюційних ходів. У еволюції найпростіших виділяють два основних філогенетичних напрямки. При першому з них, морфофізіологічні перетворення йшли у фізичних межах однієї клітини шляхом поліплоїдії та диференціювання внутрішньоклітинних структур. Найбільшої складності при даному засобі еволюції досягли інфузорії. Проте цей філогенетичний напрямок через поліплоїдію клітинних органоїдів виявився недосконалим, тому що внутрішньоклітинні структури мають обмежену спроможність до модифікацій у силу їх вже вузької спеціалізації.
Другий філогенетичний напрямок йшов через колоніальну багатоклітинність і диференціювання клітин, що їх утворюють. Даний ланцюг еволюційних подій призвів до появи багатоклітинних тварин. Подібну філогенію можна уявити і для багатоклітинних грибів і рослин.
Дати пояснення еволюційного значення кожного напрямку біологічного прогресу.
Питання для самоКОНТРОЛЮ:
1. Які докази ендосимбіотичної теорії еволюції клітин еукаріот?
2. Який структурно-функціональний внесок кожного ендосимбіонта?
3. Які структурно-функціональні умови еволюції царства Грибів, Тварин та Рослин?
4. Які обмеження прогресивної еволюції прокаріот?
5. На яких морфофункціональних особливостях заснована прогресивна еволюція еукаріот?
6. У чому причина «глухого кута» еволюції типу Найпростіших?
ТЕМА 4
БІОЛОГІЧНІ МЕМБРАНИ
МЕТА ЗАНЯТТЯ: Вивчити властивості і регуляцію біомембран – одного з обов’язкових елементів живих організмів.
Питання для самопідготовкИ:
1. Біомембрани – як обов’язковий структурний елемент клітини в передбіологічний (хімічний) період її еволюції. Загальний план будови.
2. Мембранні ліпіди: фосфоліпіди (фосфогліцероліпіди, фосфосфінголіпіди), гліколіпіди, холестерин; склад, амфіфільність.
3. Властивості біомембран: 1) утворення бішарів і їх спроможність до самозамикання; 2) рухливість, текучість бішарів; 3) асиметрія бішарів; 4) вибіркова проникність.
4. Мембранні білки.
5. Плазматична мембрана. Особливість загального плану будови.
6. Функції плазматичної мембрани: механічна, транспортна.
Навчальні завдання
ЗАВДАННЯ 1. Транспорт речовин через мембрани.
Поставте в штатив 3 пробірки (хімічні). Налийте в них по 2 мл розчину хлористого натрію різної концентрації: 0,5%, 0,85%, 5%. Додайте в кожну з них 0,05 мл (1 крапля) суцільної крові. Через 5 хвилин приготуйте тимчасовий препарат із суспензії клітин і їх залишків із кожної пробірки і мікроскопуйте під збільшенням об. 100х; ок. 10х.
На препараті з першої пробірки під мікроскопом видно залишки еритроцитів у вигляді оболонок; на другому препараті еритроцити мають нормальний вигляд; на третьому – еритроцити зморщені, з шипами.
Замалюйте по одному еритроциту з кожного препарату. Під кожним малюнком зробіть висновок про тип розчинів у досліді (гіпер-, гіпо-, ізотонічний), цитологічне явище, що відбувається з еритроцитами (плазмоліз, гемоліз, нормальна фізіологія), засоби транспорту речовин через мембрану еритроцитів (проста дифузія, полегшена дифузія, активний транспорт).
ЗАВДАННЯ 2. Фагоцитоз нейтрофілів.
Препарат виготовлено із суміші стабілізованої антикоагулянтом периферичної крові та суспензії однодобової культури стафілококу штаму 209. Після 30 хвилин інкубації із суміші готували мазок, фіксували його метиловим спиртом та забарвлювали фарбою Романовського-Гімза.
Під імерсією знайти вільні нейтрофіли та нейтрофіли, які фагоцитували мікробів. Мікроби забарвлюються у темно-вишневий колір, округлі за формою, добре контуруються. Зверніть увагу на різну кількість грудочок у фагосомах нейтрофілів. Замалювати вільний нейтрофіл та нейтрофіл, що фагоцитував. Підписати: ядро, мікроорганізм, кількість мікробних тіл. Вказати стадії фагоцитозу та його тип (специфічний, неспецифічний).
ЗАВДАННЯ 3. Утворення та рециркуляція мембран плазмолеми та вакуолярної системи (схема).
Плазмолема та вакуолярна система клітин відносяться до одномембранних структур. Вони структурно і функціонально тісно пов’язані між собою. Мембрани цих утворень разом із їх вмістом послідовно переходять одна в одну, здійснюючи спрямований потік речовин у клітині. Так, речовини, що синтезуються на мембранах ендоплазматичної сітки, пакуються у одномембранні контейнери (пухирці), які транспортуються в апарат Гольджі та зливаються з його проксимальними мембранами. Далі утворені речовини у вигляді секреторних пухирців екзоцитуються за межі клітини, зливаючись із цитолемою або перерозподіляються в інші ділянки клітини: лізосоми, пероксисоми, цитозоль. Окрім того, мембрана клітин постійно поглинається в клітину у вигляді ендосом у процесі її ендоцитозу (піно- та фагоцитозу). Ендосоми потім зливаються з первинними лізосомами та перетворюються у вторинні лізосоми. Однак поряд із вище описаним однонаправленим потоком речовин, запакованих в одномембранні везикули, існує зворотній потік пустотільних сплющених везикул, що повертають мембранам попередню структуру. Безумовно, частка цих контейнерів під час потоку речовин руйнується, наприклад, у лізосомах. Ця невелика втрата мембран поповнюється новим синтезом їх компонентів (ліпідів та білків) у гранулярному ендоплазматичному ретикулумі. Він є джерелом усіх клітинних мембран, за винятком двомембранних органел: мітохондрій і пластид. Під час прямого та зворотного потоку мембран іде їх сортування, комплектування так, що кожна цитоплазматична структура має свій специфічний набір мембран.
Замалювати основні шляхи прямого та зворотного потоків мембран у клітині з підписом відповідних органел.
Питання для самоКОНТРОЛЮ:
1. На чому ґрунтуються властивості біомембран: текучість, рухливість, здатність до самозамикання, утворення бішарів?
2. Визначити джерела енергії транспорту речовин через мембрану: проста дифузія, полегшена дифузія, активний транспорт.
3. Який принцип будови білків визначає їх властивості?
4. Яка електонно-мікроскопічна картина сприяла помилковому уявленню про будову біомембран по типу подвійного «сандвіча» і які нові методичні засоби дозволили висунути правильну «мозаїчну теорію»?
5. Вказати структурну відмінність цитоплазматичної мембрани від інших клітинних мембран.
6. Де утворюються мембрани і який їх потік у клітині?
ТЕМА 5
МІЖКЛІТИННІ КОНТАКТИ. СПЕЦІАЛІЗОВАНІ СТРУКТУРИ ПЛАЗМАТИЧНОЇ МЕМБРАНИ. КЛІТИННІ ОБОЛОНКИ
МЕТА ЗАНЯТТЯ: Вивчити типи міжклітинних контактів, ознайомитись із спеціалізованими структурами плазматичної мембрани, вивчити особливості будови оболонок клітин про- та еукаріот.
Питання для самопідготовкИ:
Рецепторна роль плазмолеми.
Міжклітинні контакти: адгезивні, замикаючі, провідні.
Похідні плазмолеми: війки, джгутики, мікроворсинки.
Оболонки клітин прокаріот, рослин, грибів, тварин.
Утворення та рециркуляція мембран.
НАВЧАЛЬНІ ЗАВДАННЯ
ЗАВДАННЯ 1. Рецепторна функція біомембран. Вивчити будову та функції рецепторних білків.
По всій плазмолемі розташовуються численні мембранні білки – рецептори. В утворенні рецепторного комплексу бере участь вуглеводний компонент глікопептидів та гліколіпідів. Рецептори беруть участь у специфічних ліганд-зв’язуючих реакціях. Це означає, що рецептор специфічно комплементарно зв’язується з одним із лігандів. Ліганд – субстрат, антигенна детермінанта, комплементарна активному центру рецептора. Частина рецепторних білків бере участь у полегшеному і активному переносі речовин через плазматичну мембрану. Через іншу частину здійснюється рецепція зовнішніх агентів і регуляція обміну речовин у клітині, її подразливість, рухова реакція та ін. Дану функцію виконують трансмембранні білки. Їх взаємодія з зовнішнім лігандом супроводжується конформаційними перетвореннями білка, що є сигналом трансмембранного переносу інформації. Наприклад, у такий спосіб гормонами регулюється активність клітин-мішеней, що мають до них рецептори. Через рецептор відбувається передача інформації від гормону до клітини. При цьому гормон не проникає в клітину. Утворення на зовнішній стороні комплексу гормону (ліганду) і рецептора (його активного центру) супроводжується конформаційними змінами рецептора. При цьому змінюється третинна структура цитоплазматичної ділянки рецептора (хвоста). Дана зміна є сигналом для активації асоційованих із рецепторами, але вже з внутрішньої сторони цитоплазматичної мембрани білків-ферментів, що генерують із АТФ і ГТФ утворення біологічно активних циклічних нуклеотидів, відповідно, циклічного аденозинмонофосфату (цАМФ) і циклічного гуанозинмонофосфату (цГМФ). У свою чергу цАМФ і цГМФ активують спеціальні ферменти кінази, що здійснюють процеси фосфорилювання – перенесення фосфатних груп від АТФ до молекул клітинних білків, нуклеїнових кислот. Фосфорилювання призводить до активації або інактивації цих біополімерів. Після стихання зовнішнього сигналу (у відсутності гормону) циклічні нуклеотиди розпадаються, а фосфатні групи знімаються з біополімерів, повертаючи їх із напруженого стану в те, що було до зовнішнього сигналу.
Подібний тип регуляції клітин-мішеней відбувається при імунних реакціях, дії нервової системи, міжклітинних контактах.
Складіть усно схему причинно-наслідкових відношень у приведеному прикладі регуляції активності біорегуляторів клітини гормоном «X», включаючи етапи: 1) взаємодія рецептора з лігандом (гормоном); 2) активації внутрішньоклітинної аденілциклази; 3) синтез цАМФ; 4) активація кіназ; 5) фосфорилювання (модифікація) біополімерів (білків, нуклеїнових кислот) зі зміною їх функціональної активності.
ЗАВДАННЯ 2. Латеральна дифузія рецепторів у плазмолемі.
Роздивіться схему взаємодії ліганду (антигена) з рецептором імуноглобулінової природи (антитіло) на поверхні В-лімфоцита. Антиген мічений флуорохромом (флуорисцеїном), тому його розташування на поверхні клітини можна реєструвати за допомогою люмінесцентного мікроскопа. На першому етапі взаємодії мітка (флуоресцеїн) розподілена рівномірно, але через деякий час мітка накопичується у вигляді окремих плям (матчінг), а потім збирається разом, утворюючи комплекс (кеппінг). Далі комплекс ліганд-рецептор піноцитується з утворенням ендосоми, в якій антиген фрагментується ферментами лізосом. Через 30 хвилин рецептори до антигена знов регенеруються на поверхні В-лімфоцита.
Схематично замалювати етапи: 1) адгезії антигена; 2) матчінг; 3) кеппінг; 4) ендоцитоз. Пояснити вказані феномени з позицій біологічних властивостей.
ЗАВДАННЯ 3. Міжклітинні контакти.
Роздивіться в практикумі по цитології (див. рекомендовану літературу) мікрофотографії, зроблені за допомогою електронного мікроскопа (мал. 140-144). Використовуючи приведені описи препаратів, зробіть аналіз структури адгезивних контактів (простий контакт, зубчастий контакт, «замок», десмосома), ізолюючого контакту (щільний контакт) і провідного контакту (щілинний контакт). Відзначте їх основні морфологічні особливості та функції.
ЗАВДАННЯ 4. Війки епітеліальних клітин кишечнику беззубки.
Одношаровий епітелій, який вистилає кишечник, складається з високих циліндричних клітин, розташованих на базальній мембрані. Ядра в цих клітинах розташовуються на різних рівнях, але завжди в базальних частинах клітин. Базальна частина клітини прилягає до базальної мембрани, апікальна звернена в кишкову порожнину і покрита війками. Ядра мають овальну форму. В них помітні глибки хроматину і ядерця. Апікальна поверхня клітин покрита близько розташованими один до одного війками. Їх синхронний рух створює невпинний потік рідини. При вивченні з імерсійною системою апікальних частин епітеліальних клітин видно, що в основі війок розташовується переривчаста лінія. Вона складається з близько розташованих один до одного базальних зерен, від кожного з яких і відходить війка. Під збільшенням об. 40х; ок. 10х замалюйте ділянку епітелію і підпишіть: 1) війки, 2) війчасті клітини, 3) ядро, 4) базальну мембрану.
ЗАВДАННЯ 5. Мікроворсинки апікальної поверхні кишкового епітелію і ниркового епітелію хребетних.
За допомогою електронних мікрофотографій, поданих у практикумі (мал. 149, 150) познайомтеся з ультраструктурою мікроворсинок. Використовуйте описи мікрофотографій і підписи до них. Мікроворсинки покриті плазмолемою із глікокаліксом. Усередині них видно мікрофіламенти.
Питання для самоКОНТРОЛЮ:
Який молекулярно-клітинний механізм матчінгу та кеппінгу комплексу ліганд-рецептор?
Як передається в цитоплазму сигнал про зв’язок ліганду з рецептором?
Чим обумовлена міцність зв’язку клітини за допомогою крапкових демосом?
Яка різниця між скелетними структурами, які складають крапкові десмосоми та опоясуючі десмосоми?
Назвати головні функції щільного контакту.
Назвати відмінні механізми проведення сполук щільним контактом і синапсом.
Назвати основні структурно-функціональні відмінності мікроворсинок і війок.
ТЕМА 6
ЯДРО КЛІТИНИ. ДНК ТА БІЛКИ ХРОМАТИНУ. РІВНІ ОРГАНІЗАЦІЇ ХРОМОСОМ
МЕТА ЗАНЯТТЯ: Вивчити будову та еволюцію генетичного апарата клітини.
Питання для самопідготовкИ:
1. Основні функції ядра. Загальний план будови.
2. Еволюційні аспекти будови генетичних структур організмів.
3. Інтерфазні хромосоми. Еу- і гетерохроматин.
4. ДНК хромосом. Фракції ДНК.
5. Особливості організації геному еукаріот.
6. Білки хроматину.
7. Рівні структурної організації хромосом.
Навчальні завдання
ЗАВДАННЯ 1. Графологічна структура молекулярних і цитологічних рівнів організації генетичного апарату клітин еукаріот.
Розгляньте і замалюйте представлену схему. Позначте молекулярні та цитологічні світлооптичні рівні аналізу генетичного апарата еукаріот.
ЗАВДАННЯ 2. Співвідношення кодуючих та не кодуючих ділянок ДНК у про- та еукаріот.
Розгляньте умовну ділянку хромосоми еукаріот і фракції ДНК, які знаходяться на ній. Визначте відсоткове співвідношення кодуючої та не кодуючої ДНК по вимірюванню лінійних розмірів. Порівняйте це співвідношення у прокаріот. Зробіть висновок про еволюційний потенціал цих двох надцарств організмів.
ЗАВДАННЯ 3. Рівні структурної організації хромосом еукаріот.
Розгляньте і замалюйте схему рівнів структурної організації хромосом еукаріот. Виділіть на кожному з них наступні ознаки: назва рівня, структурна одиниця, поперечний діаметр, ступінь скорочення ДНК, участь кислих та лужних білків, нерівномірність укладки та чим вона обумовлена.
1-й рівень конденсації хроматину – нуклеосомний. Структурною одиницею є нуклеосома – гістоновий октамер шароподібної форми (за іншими даними у вигляді шайби). Вона утворена двома молекулами гістонів кожної фракції (Н3, Н2а, Н2б, Н4). Навколо нуклеосоми закручена молекула ДНК. Ділянка ДНК між нуклеосомами називається лінкерною. Лінкерні ділянки ДНК асоційовані з Н1 гістоном, який зближує нуклеосоми в одну нитку діаметром 10 нм (діаметр нуклеосом). Перший рівень укладки дає скорочення в 7 разів. Нуклеосомна організація хромосом є функціонально активною: гени, що помірно транскрибуються, зберігають нуклеосомну організацію. При інтенсивній транскрипції (наприклад, рибосомальні гени) ДНК звільняється від нуклеосом.
2-й рівень укладки – нуклеомерний. Структурною одиницею є нуклеомер. Нуклеосомна нитка спіралізується навколо уявної осі, утворюючи нитку другого порядку діаметром 20 або 30 нм. Існує дві моделі укладки хроматину цього рівня:
1) соленоїдна – рівномірна спіралізація у вигляді індуктивної котушки (соленоїд);
2) глобулярна (найбільш достовірна) – спіралізація ниток ДНП відбувається нерівномірно у вигляді глобул, які об’єднують 8-10 нуклеосом. Утворена одиниця називається нуклеомером. В утворенні нуклеомерів істотну роль відіграє гістон Н1, однак не виключена участь кислих білків та гетерохроматинових ділянок ДНК. Нуклеомерна укладка дає скорочення в 6 разів (загальне скорочення – понад 40 разів).
3-й рівень укладки – доменний, хромомерний, хромонемний (кожен із цих термінів описує одну з ознак укладки ДНП). Структурна одиниця – хромомер. Даний рівень найменш вивчений і може включати ще кілька проміжних етапів.
Перший етап: нитка ДНП другого порядку утворює петлі, які сходяться в одній точці (зближуються в центрі) і скріплюються кінцями кислих білків до осьової структури хромосоми і до не гістонового матриксу інтерфазного ядра. Утворюється петлеподібна розетка – хромомера. Довжина петель – 10-30 нм (у середньому 3000 н.п.), кількість петель 15-20 шт. У місцях прикріплення петель до білкової осьової структури хромосоми та білкового ядерного матриксу розташовані реплікаційні центри, з яких починається реплікація молекул ДНК у S-періоді. Ймовірно, одна петля є одиницею реплікації (репліконом). Хромосоми еукаріот починають реплікуватись одразу в багатьох місцях, пов’язаних із петлями розеток, тому за цією ознакою хромосоми еукаріот є полірепліконними.
Другий етап третього рівня: компактизація петель розетки (хромомери). Припускають, що кожна з петель розетки спіралізується навколо своєї осі, внаслідок чого утворюється петлеподібна (спіралізована) розетка діаметром 0,2-0,3 мкм (діаметр хромомер). У цей час хромосома представляє собою сукупність таких хромомер, накладених одна на одну, та з’єднаних міжхромомерними лінкерними ділянками ДНП.
Третій етап цього рівня: зближення хромомер за допомогою кислих білків, у результаті чого утворюється нитка – хромонема – діаметром 0,2-0,3 мкм.
Четвертий рівень конденсації хроматину – хромосоми клітин, що діляться. Вважають, що починаючи з профази стопки хромомер, що складають хромонему, ще більше наближаються одна до одної за рахунок кислих білків із одночасною спіралізацією навколо осі. Залежно від своєї довжини, хромосома робить від 4 до 10 великих витків. Як наслідок цього утворюється структура метафазної хромосоми діаметром 1,0-1,5 мкм. Укорочення ДНК цього рівня складає 8-10 разів, а з урахуванням усіх рівнів кінцеве укорочення ДНК складає понад 104 разів. Даний рівень укладки хроматину, як і на попередніх рівнях, також не рівномірний. Ця не рівномірність конденсації проявляється у вигляді хромодисків по довжині хромосом у клітинах, що діляться. Хромодиск є структурною одиницею метафазної хромосоми. Він представляє собою скупченість конденсованого хроматину. Величина і розташування дисків приблизно подібна для гомологічних хромосом і специфічна для різних пар. Це дає можливість складати каріотип індивіду за парами гомологічних хромосом.
Дати інтерпретацію причин унікальної нерівномірної конденсації ДНК не гомологічних хромосом на кожному рівні і як ця ознака використовується при каріотипуванні.
Питання для самоКОНТРОЛЮ:
1. Назвати загальні структурно-генетичні ознаки всіх живих організмів Землі.
2. Назвати специфічні особливості будови генетичного апарата про- та еукаріот.
3. Які структури хромосом ми розрізняємо в світовому мікроскопі?
4. Розкрийте поняття «функціонально активний еухроматин». Вказати його молекулярно-генетичний структурний рівень.
5. Чому видове різноманіття кислих білків значне, тоді як гістони за цією ознакою консервативні?
6. Чим обумовлена нерівномірність укладки хромосом на кожному з її рівнів і як ця ознака використовується в каріотипуванні?
ТЕМА 7
ЯДРО КЛІТИНИ. МЕТАФАЗНІ ХРОМОСОМИ. СТАТЕВИЙ ХРОМАТИН. КАРІОТИП
МЕТА ЗАНЯТТЯ: Вивчити будову метафазних та анафазних хромосом, ознайомитись із методами вивчення структури хромосом за типом диференційного забарвлення.
Питання для самопідготовкИ:
1. Будова метафазних та анафазних хромосом.
2. Типи диференційного забарвлення хромосом за методами Q, G, C, Ag-дисків.
3. Структурна та генетична характеристика сестринських та гомологічних хромосом.
4. Поняття каріотипу. Типи плоїдності ядра.
5. Статевий хроматин Х та У. Природа його утворення.
НАВЧАЛЬНІ ЗАВДАННЯ
ЗАВДАННЯ 1. Порівняння каріотипів різних видів.
Під імерсійним мікроскопом порівняйте каріотипи миші, пацюка, людини. Зверніть увагу на розходження числа, розмірів і форми хромосом. Зазначте число хромосом кожного проаналізованого виду. У миші в каріотипі 40 хромосом, усі вони акроцентричного типу; у пацюка – 42 хромосоми, більша частина з них – акроцентричні, інші – субметацентричні; у клітинах людини 46 хромосом: мета-, субмета- і акроцентричного типу.
ЗАВДАННЯ 2. Типи хромосом.
На мікрофотографіях метафазної пластинки людини знайдіть 3 типи хромосом: метацентричну, субметацентричну та акроцентричну. Схематично замалюйте по одній хромосомі кожного типу, відмітивши на одній із них деталі будови: хроматида, плече хроматиди, центромера, вторинна перетяжка, супутник.
ЗАВДАННЯ 3. G-диски і С-диски при диференційне фарбуванні хромосом.
На мікрофотографії метафазної пластинки хромосом людини (диференційно пофарбовані G-диски – Гімза-диски) роздивіться поперечну посмугованість хромосом. Зверніть увагу, що малюнок цієї посмугованості однаковий для гомологічних хромосом і різний для не гомологічних хромосом. Поперечна посмугованість обумовлена специфічним для кожної пари гомологічних хромосом розподілом по довжині ділянок еу- і гетерохроматину, що у свою чергу визначає особливість нерівномірної конденсації хроматину при підготовці хромосом до поділу.
На іншій мікрофотографії метафазної пластинки хромосоми пофарбовані по методу, що виявляє прицентромерний гетерохроматин (С-диски). Зверніть увагу на великі диски (С-диски) у 1, 9 і 16 хромосомах, а також на гетерогенність С-дисків у 1 парі гомологічних хромосом усього набору. Зробіть висновок про природу С-дисків та причини їх відмінностей у материнських і батьківських хромосомах. Гетерохроматин, у тому числі і прицентромерний, не містить генів і представлений фракцією високо повторюваних нуклеотидних послідовностей ДНК. Вони несуть в основному структурну функцію, а також беруть участь у регуляції активності генів еухроматичних районів.
ЗАВДАННЯ 4. Статевий хроматин Х у клітинах ротової порожнини людини.
Стерильним ватним тампоном протріть слизову поверхню щоки для зняття клітин із пікнотичними ядрами. Стерильним шпателем сильним рухом зробіть зіскоб зі слизової поверхні щоки. З отриманого матеріалу рухом в один бік зробіть мазок.
Додайте до нього 2 краплі 1% ацетоорсеїну і накрийте покривним склом. Через 2 хвилини обережно зверху промокніть препарат для видалення фарби з мазків. Мікроскопуйте під імерсією. Аналізуйте клітини проміжного шару. Зверніть увагу на морфологію ядра, пристінкове положення великих глибок хроматину. У частини клітин жіночої статі знайдіть пристінкові глибки хроматину розміром 1-3 мкм (тільце Барра). Це Х-хроматин. Замалюйте цю клітину. Зробіть висновок про природу Х-хроматину і набору статевих хромосом у даного індивіда.
Х-хроматин являє собою конденсовану Х-хромосому. Інша Х-хромосома в жіночому наборі і єдина Х-хромосома в чоловічому наборі знаходяться, як і всі інші хромосоми, в деконденсованому, генетично активному стані. Інактивація однієї з X-хромосом відбувається на ранніх етапах дроблення зиготи. Причому в одних бластомерах інактивується Х-хромосома чоловічого походження, в інших – жіночого. Біологічне значення інактивації однієї з Х-хромосом полягає у зрівнянні дози активних генів, розташованих у Х-хромосомі у осіб чоловічої та жіночої статі. Додаткові Х-хромосоми за рахунок мутацій також інактивуються і генетично активною залишається тільки одна Х-хромосома. Статевий хроматин по типу Х-хроматину характерний для всіх видів тварин із гетерогаметною статтю.
ЗАВДАННЯ 5. Аналіз каріотипу людини за Денверською номенклатурою.
Візьміть із великого пакета фотографію метафазної пластинки і маленький пакетик із вирізаними хромосомами з цієї фотографії. Перенесіть хромосоми з пакетика на чистий лист паперу і переверніть їх лицем догори, а потім перерахуйте. Їх кількість повинна збігатися з числом зображених на мікрофотографії. Хромосоми на поданих мікрофотографіях пофарбовані рутинним способом (рівномірне забарвлення по всій довжині), що не дозволяє в більшості випадків виділяти гомологічні хромосоми. Для цього необхідно диференційне забарвлення (див. завдання 3 заняття 6). При рутинному фарбуванні хромосоми можна класифікувати тільки по групах, за винятком груп А та Е, де можна виділити гомологічні хромосоми, використовуючи центромірний індекс.
Денверська номенклатура хромосом людини
|
Групи хромосом
1 |
Номер і центральний індекс пари гомологічних хромосом 2 |
Морфологія хромосом
3 |
|
А |
1(49), 2(38), 3(47). |
1.Найбільша метацентрична 2.Найбільша метацентрична 3.Велика метацентрична |
|
В |
4; 5(29) |
Великі субметацентричні хромосоми (центромера сильно зсунута до кінця) |
|
С |
6(37),-Х- 12(29) |
Середні субметацентричні |
|
D |
13,14,15(18) |
Акроцентричні великі. Короткі плечі є вторинними перетяжками, що часто мають супутники |
|
Е |
16(40),17(34),18(29) |
Маленькі субметацентричні |
|
F |
19; 20(46) |
Маленькі метацентричні |
|
G та Y |
21; 22(18) |
Маленькі акроцентричні. Короткі плечі є вторинними перетяжками, що часто мають супутники (окрім Y-хромосоми) |
Примітка: У дужках поданий центромірний індекс (ЦІ) у відсотках, розрахований за формулою:
ЦІ = ДОВЖИНА КОРОТКОГО ПЛЕЧА × 100
ДОВЖИНА ВСІЄЇ ХРОМОСОМИ
За допомогою опису морфології хромосом, складіть каріотип людини, розкладаючи хромосоми в рядках відповідних груп таблиці Денверської номенклатури, записаної в альбомі: на початку знайдіть 1, 2, 3 пари хромосом групи А, потім вибирайте за зменшенням розміру хромосоми групи B, D, С, Y (якщо вона є), F, Е. У групі Е при ретельному аналізі можна виділити гомологічні пари. Останніми вивчайте хромосоми групи С. В ній повинно в нормі утримуватися 18 хромосом при жіночому каріотипі (16 аутосом 6-12 пар і дві хромосоми), при чоловічому 17 (16 аутосом і 1 Х-хромосома). У жіночому каріотипі маленьких акроцентричних хромосом 4 (21-22 пари), у чоловічому – 5 (21-22 пари та Y-хромосома). Зробіть висновок про відповідність числа хромосом нормальному каріотипу людини і визначте його стать.
Питання для самоКОНТРОЛЮ:
1. Загальні риси та відмінності в будові хромосом про- та еукаріот.
2. Надлишковість геному еукаріот – плата за еволюцію?
3. Які структурно-генетичні ознаки забезпечують різну укладку хромосом на всіх рівнях їх організації?
4. Яка природа Х та У хроматину?
5. Де шукати С хроматин в інтерфазних ядрах?
6. Які порушення генотипу можна встановити при аналізі каріотипу при рутинному та диференційному забарвленні хромосом?
ТЕМА 8
ЯДРО КЛІТИНИ. НЕ ГЕНЕТИЧНІ СТРУКТУРИ. ОБОЛОНКА, КАРІОПЛАЗМА, МАТРИКС. ЯДЕРЦЕ ТА РИБОСОМИ
МЕТА ЗАНЯТТЯ: Вивчити динамічні взаємовідносини компонентів ядра та їх зв’язок із цитоплазмою.
Питання для самопідготовкИ:
1. Будова і функції ядерно-цитоплазматичної оболонки.
2. Поняття про ядерно-цитоплазматичне співвідношення.
3. Будова і функції ядерця.
4. Генетика ядерцевого організатора.
5. Рибосоми про- та еукаріот.
6. Ядерний матрикс, ядерний сік.
7. Динаміка ядерця в мітотичному циклі.
Навчальні завдання
ЗАВДАННЯ 1. Порівняння структурних особливостей рибосом про- та еукаріот.
Перевірте порівняння структурних особливостей рибосом про- та еукаріот.
|
Тип надцарства |
Тип рибо-сом, S |
Локалізація в клітині |
Субодиниця S |
Тип рРНК у субодиниць |
Число кислих білків у субодиницях рибосом |
|
|
|
Прокаріоти |
70 |
Цитоплазма |
30; 50 |
16;23;5 |
55 |
| |
|
Еукаріоти |
80 70 |
Цитоплазма Мітохондрії, пластиди |
40; 60 - |
18;28;5;5,8 - |
100 - |
|
ЗАВДАННЯ 2. Організація і функції ядерцевого організатора у Xenopus levis.
Проаналізуйте та замалюйте цистрон ядерцевого організатора (ЯО), підпишіть його структуру. Потім представте потік реалізації інформації у вигляді малюнка про-рРНК до зрілих типів рРНК. При цьому вкажіть тип РНК-полімерази, одиниці седиментації про-рРНК та зрілих рРНК, ферменти процесингу, відповідність усіх видів нуклеїнових кислот цитологічним компонентам ядерця.
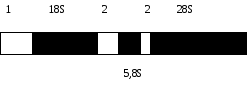
де 1 і 2 – транскрибовані послідовності ДНК (зовнішня і дві внутрішні);
18s, 28s та 5,8s – ділянки, що відповідають зрілим рибосомальним РНК.
ЗАВДАННЯ 3. Ядерцевий матеріал на мітотичних хромосомах людини.
У вторинних перетяжках коротких плечей акроцентричних хромосом людини тандемно розташовуються до 50 копій рибосомальних цистронів, що кодують 18S, 5,8S, 28S рРНК. Це ядерцеутворюючі райони (ЯУР), які функціонують в інтерфазі. У період вступу клітини до мітозу активність ЯУР припиняється, матеріал ядерця резорбується (переходить у цитоплазму), але не цілком. Частина ядерцевого матеріалу залишається пов’язаною з ЯУР. Залишковий матеріал складається з кислих білків, що включають регуляторні білки, РНК-полімеразу. Ці білки вибірково фарбуються азотно-кислим сріблом у вигляді чорних гранул. Скупчення їх в області ЯУР називають Ag-дисками. Роздивіться мікрофотографію метафазної пластинки людини після фарбування азотно-кислим сріблом. За допомогою цієї фотографії можна простежити зв’язок будови хромосом із їх функцією. Зверніть увагу на те, що не всі 10 акроцентричних хромосом мають Ag-диски. Окрім того, розмір наявних Ag-дисків не однаковий у різних хромосом. Доведено, що азотно-кислим сріблом забарвлюються тільки ті ЯУР, рибосомальні гени яких були активними в попередній інтерфазі. Від ступеня активності рибосомальних генів залежить розмір Ag-дисків. Замалюйте на вибір одну акроцентричну хромосому з неактивним ЯУР і одну з активним ЯУР, що несе Ag-диски, зробивши відповідні позначення: активний або неактивний ЯУР, Ag-диски, акроцентрична хромосома, коротке плече, довге плече, вторинна перетяжка. Підкреслюючи значимість цієї роботи, зробіть висновок, що неоднакова активність генів характерна не лише для генів рРНК, але і для унікальних генів, що кодують специфічні білки. З вибірковою активністю унікальних генів пов’язане диференціювання соматичних клітин, що утворюють тканини та органи.
ЗАВДАННЯ 4. Шляхи транспорту речовин між ядром та цитоплазмою.
Існує декілька шляхів транспорту речовин із ядра в цитоплазму і навпаки.
1) транспорт через ядерні мембрани
1а) транспорт через внутрішню ядерну мембрану з переходом у цистерни ЕС через вирости у зовнішній ядерній мембрані. Таким чином відбувається обмін іонів та мікромолекул (моносахаридів, нуклеотидів, амінокислот);
2) інвагінація внутрішньої ядерної мембрани з утворенням пухирця, що переходить далі в цистерни ендоплазматичної сітки (ЕС); або
2а) безпосередньо у цитозоль після злиття пухирця із зовнішньою ядерною мембраною. Так можуть дифундувати великі макромолекули (білки, РНК)
3) транспорт через внутрішню мембрану з утворенням та відокремленням пухирця в зовнішній мембрані. Набір речовин, що переноситься, аналогічний першому способу перенесення, хіба що більш активний.
4) транспорт через порові комплекси (ПК) – основний вид транспорту всіх мікро- і макромолекул. При цьому транспорт РНК через ПК супроводжується процесингом. Цей процес активний і йде за участю рецепторних білків, ферментів, АТФ.
5) вип’ячування ділянки ядерної оболонки на всю довжину з відщепленням утворених виростів.
Способи потоку речовин із ядра в цитоплазму та з цитоплазми в ядро за участю ядерної оболонки подібні. Слід відмітити, що перенесення речовин із ядра в цитоплазму шляхом відщеплення мембран ядра є також свідченням рециркулюючого потоку мембран у клітині. Так, відщеплення везикул із ядерної мембрани припускає їх вставлення, але вже за рахунок везикул ендоплазматичної сітки, апарата Гольджі, плазмолеми.
6) ядерно-цитоплазматичний обмін також має місце при поділі клітин, коли руйнується ядерна оболонка.
Роздивіться запропоновану схему однонаправленого потоку речовин із ядра в цитоплазму. Замалюйте її та додайте зворотній потік речовин у ядро з практикуму. Вкажіть на схемі цифрами тип потоку, а в тексті під малюнком дайте розшифровку даних цифр із переліком речовин, що переносяться.
Питання для самоКОНТРОЛЮ:
1. Назвати загальні ознаки для зовнішньої мембрани і мембрани ендоплазматичної сітки.
2. Перелічити основні структурні елементи порового комплексу.
3. Вказати механізми транспорту речовин через поровий комплекс.
4. Зробити структурно-біохімічне порівняння ядерця.
5. Де синтезується 5 S рРНК у клітинах людини?
6. Чим пояснити наявність 70 S рибосом у мітохондріях і пластидах?
7. Назвати основні компоненти ядерного матриксу.
ТЕМА 9
ОРГАНЕЛИ ЦИТОПЛАЗМИ. ГІАЛОПЛАЗМА. ВАКУОЛЯРНА СИСТЕМА
МЕТА ЗАНЯТТЯ: Вивчити систему одномембранних структур, пов’язаних між собою спільністю структури, функції і походженням.
Питання для самопідготовкИ:
1. Гіалоплазма. Структура, функції, участь у проміжному обміні.
2. Поняття про вакуолярну систему цитоплазми. Ендоплазматична сітка.
3. Різноманіття структури і функцій апарата Гольджі.
4. Лізосоми. Походження, розвиток, участь у гетеро- і аутофагії.
5. Походження і рециркуляція мембран клітини.
Навчальні завдання
ЗАВДАННЯ 1. Гранулярна ендоплазматична сітка.
Роздивіться електронні мікрофотографії та описи до них, приведені в практикумі по цитології:
Препарат № 51 – електронна мікрофотографія ділянки клітини шовковидільної залози гусениці тутового шовкопряда.
Препарат № 55 – електронна мікрофотографія плазматичної клітини.
Препарат № 57 – електронна мікрофотографія тигроїда (тільце Нісля) у нейроні.
Зверніть увагу на залежність ступеня розвитку гранулярної ендоплазматичної сітки від інтенсивності синтезу секреторних білків (фіброїну) в шовковичній залозі шовкопряда та імуноглобуліну в плазматичній клітині. Концентрично розташовані мембрани з рибосомами щільно прилягають одна до одної. Розташовані між ними канали заповнені синтезованим продуктом. Гранулярна ендоплазматична сітка займає великі ділянки цитоплазми. Секретовані в ній білки за допомогою апарата Гольджі виводяться за межі клітини.
Тигроїд або тільце Нісля в нейронах – це скупчення щільно упакованих мембран ендоплазматичної сітки у вигляді окремих ділянок у цитоплазмі. Кількість тигроїду прямо корелює з інтенсивністю синтезу білків у нервових клітинах, що використовуються в основному на відновлення мембранних структур клітини.
ЗАВДАННЯ 2. Базофілія цитоплазми трансформованих лімфоцитів у культурі
клітин із фітогемаглютиніном.
Фітогемаглютинін – лектин із квасолі посівної, є поліклональним мітогеном для Т-лімфоцитів. При культивуванні з ним лімфоцитів більшість Т-лімфоцитів трансформується в бластні клітини з наступною проліферацією. З культури лімфоцитів, стимульованих фітогемаглютиніном, виготовлений мазок і пофарбований азур-еозином за Романовським-Гімза. У мазку зустрічаються нестимульовані малі лімфоцити і лімфоцити, трансформовані в бластні клітини (великі і малі бласти). Це великі клітини 12-18 мкм і більше. Ядро містить деконденсований хроматин, добре розвинені одне або декілька ядерець. Цитоплазма широка, з різкою базофілією, особливо по периферії клітини. Базофілія цитоплазми обумовлена розвитком системи, що синтезує білок – гранулярної ендоплазматичної сітки, яка містить велику кількість рРНК, іРНК (вони забарвлюються в синій колір азуром, що має основні властивості).
Замалюйте по одному малому лімфоциту і бласттрансформованому лімфоциту, зробіть підпис до малюнка: 1) ядро, 2) ядерце, 3) базофільна цитоплазма.
ЗАВДАННЯ 3. Гладка ендоплазматична сітка.
Зробіть аналіз електронних мікрофотографій і описів до них, поданих у практикумі по цитології:
Препарат № 64 – гладка ендоплазматична сітка в інтерстиціальних клітинах сім'яника морської свинки.
Препарат № 67 – гладка ендоплазматична сітка в облямівкових клітинах фундальних залоз дна шлунка пацюка.
Препарат № 68 – гладка ендоплазматична сітка в клітинах печінки.
Майже вся цитоплазма цих клітин заповнена мембранами ендоплазматичної сітки без рибосом. Велика кількість гладкого ретикулуму в інтерстиціальних клітинах сім’яника обумовлена виділенням цими клітинами великої кількості стероїдного гормону, а розвинений ретикулум облямівкових клітин шлунка пов’язаний із виділенням ними хлоридів, із яких у порожнині шлунка утворюється соляна кислота. Агранулярний ретикулум клітин печінки містить ферменти, які відповідають за обмін глікогену (його синтез і розпад).
ЗАВДАННЯ 4. Апарат Гольджі у нервових клітинах спинального ганглія кошеняти.
У спинальних гангліях або спинномозкових вузлах, розташованих по ходу задніх корінців спинного мозку, локалізуються чутливі нервові клітини. Вони являють собою перший нейрон рефлекторної дуги. Ганглій зовні покритий сполучнотканинною капсулою, від якої усередину вузла проникають тонкі прошарки сполучної тканини. Великі нейрони розташовуються в основному групами в периферичних відділах. У результаті фіксації та імпрегнації осмієвою кислотою при малому збільшенні мікроскопа видно, що нервові клітини виглядають по-різному. Деякі з них суцільно пофарбовані в чорний колір і апарат Гольджі видно у них дуже погано. На таких клітинах не варто зупиняти увагу. Треба знайти нейрони, в яких уже при невеликому збільшенні було б видно ядро, межі клітини і світлу цитоплазму. На фоні світлої цитоплазми таких клітин видно значні чорні «ніші». На препараті при роботі з імерсійним об’єктивом видно: на світлому фоні цитоплазми виділяється петляста сітка, що локалізується навколо ядра. Вона складається з вигнутих і анастомозуючих між собою ниток і щаблин. У інших клітинах апарат Гольджі не утворює суцільної сітки, а складається з окремих паличок, лусочок, фрагментів різноманітної форми, не пов’язаних між собою. Варто звернути увагу на форму клітин, на світлі, майже безструктурні ядра, на світлому фоні яких чітко видно ядерця сіро-жовтого кольору. Відростків клітин при даному засобі опрацювання тканини може бути не видно.
Замалюйте 2-3 клітини, підпишіть: 1) ядро, 2) ядерце, 3) цитоплазма, 4) диктіосоми апарата Гольджі.
ЗАВДАННЯ 5. Жирові включення в клітинах печінки аксолотлю.
Жир у тій або іншій кількості може накопичуватися в різноманітних клітинах тваринного організму, у тому числі і клітинах печінки. При опрацюванні тканини чотириокисом осмію з наступним дофарбуванням зрізів карміном (ядерним барвником), видно, що в гіалоплазмі гепатоцитів локалізуються чорні жирові краплі, що адсорбували осмій. Ці краплі можуть бути різного розміру, кількість їх теж варіює. Спочатку синтезований у клітинах жир відкладається в них у вигляді дрібних крапель. Ці краплі, зливаючись між собою, утворюють краплі великого розміру. Замалюйте 2-3 клітини печінки. Зробіть підпис: 1) краплі жиру, 2) ядро клітини.
Питання для самоКОНТРОЛЮ:
1. Яка цитологічна картина гіалоплазми в стані золю та гелю відповідно?
2. Які речовини синтезуються в гіалоплазмі?
3. Вказати загальні ознаки та відмінності в будові апарата Гольджі клітин рослин та тварин.
4. Визначте біологічне значення ауто- та гетерофагії, здійснюваних у лізосомах.
5. Знайти цитологічне підтвердження вислову лікарів: «Омолодження і лікування через дозоване голодування».
6. Чим обумовлена базофілія цитоплазми метаболічно активних клітин?