

Функции и свойства скелетных мышц
Скелетная мускулатура является составной частью опорно-двигательного аппарата человека. При этом мышцы выполняют следующие функции:
1)обеспечивают определенную позу тела человека;
2)перемещают тело в пространстве;
3)перемещают отдельные части тела относительно друг друга;
4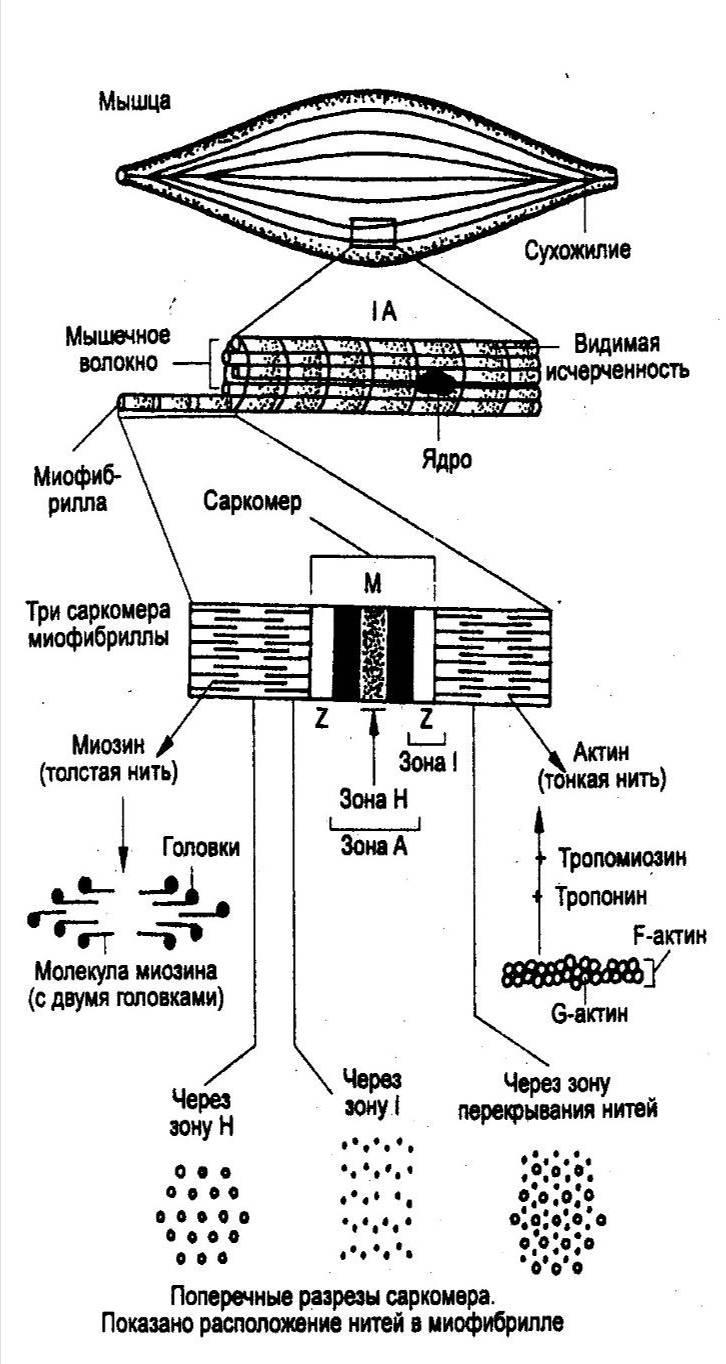 )являются источником тепла, выполняя
терморегуляционную функцию.
)являются источником тепла, выполняя
терморегуляционную функцию.
Скелетная мышца обладает следующими важнейшими свойствами:
1)возбудимостью —способностью отвечать на действие раздражителя изменением ионной проводимости и мембранного потенциала. В естественных условиях этим раздражителем является медиатор ацетилхолин, который выделяется в пресинаптических окончаниях аксонов мотонейронов. В лабораторных условиях часто используют электрическую стимуляцию мышцы. При электрической стимуляции мышцы первоначально возбуждаются нервные волокна, которые выделяют ацетилхолин, т. е. в данном случае наблюдается непрямое раздражение мышцы. Это обусловлено тем, что возбудимость нервных волокон выше мышечных. Для прямого раздражения мышцы необходимо применять миорелаксанты — вещества, блокирующие передачу нервного импульса через нервно-мышечный синапс;
2)проводимостью —способностью проводить потенциал действия по мембране вдоль и в глубь мышечного волокна по Т-трубочкам;
3)сократимостью —способностью укорачиваться или развивать напряжение при возбуждении;
4)эластичностью —способностью развивать напряжение при растягивании.
Структурная организация мышечного волокна
Мышечное волокно - это многоядерная структура, окружена мембраной и содержит сократительный аппарат — миофибриллы.В саркоплазме клеток содержатсямитохондрии,системы продольных трубочек —саркоплазматическая сеть(ретикулум) и система поперечных трубочек —Т-система.Функциональной сократительной единицей мышечной клетки является саркомер (рис. 17); из саркомеров состоит миофибрилла. Саркомеры отделяются друг от друга Z-пластинками. Они последовательно расположены в миофибрилле, поэтому сокращение саркомеров вызывает сокращение миофибриллы и общее укорочение мышечного волокна.
При изучении мышечных волокон в световой микроскоп выявило их поперечную исчерченность. Более тонкое, электронно-микроскопическое исследование показало, что поперечная исчерченность связана с особой организацией двух сократительных белков миофибрилл — актина(молекулярная масса 42 000) имиозина(молекулярная масса около 500000).
Актиновые филаменты представлены двойной нитью, закрученной в двойную спираль с шагом около 36,5 нм. Их длина 1 мкм и диаметр 6—8 нм, одним концом прикреплены к Z-пластинке, количество актиновых нитей достигает около 2000.
Между нитями актина в центре саркомера располагаются толстые нити миозина длиной около 1,6 мкм. В световом микроскопе это выглядит как полоска темного цвета (вследствие двойного лучепреломления) — анизотропный А-диск. В середине его видна более светлая полоска Н, в ней в состоянии покоя нет актиновых нитей. По обе стороны темного диска видны светлые изотропные полоски — I-диски, образованные нитями актина. В состоянии покоя нити актина и миозина незначительно перекрывают друг друга таким образом, что общая длина саркомера составляет около 2,5 мкм. Нити миозина в состоянии покоя удерживает М-линия, которая располагается в центре Н-полоски.
На боковых сторонах миозиновых нитей были обнаружены поперечные мостики (рис.18). Они состоят из головки и шейки. Головка представляет собой шарнир и может поворачиваться вокруг своей оси за счет эластических свойств. Головка при связывании с актином приобретает выраженную АТФ-азную активность.
На поперечном срезе мышечного волокна можно увидеть гексагональную организацию миофиламента: каждая нить миозина окружена шестью нитями актина (рис. 17).
В продольных бороздках актиновых филаментов располагаются нитевидные молекулы белка тропомиозина.С шагом, равным 40 нм, к молекуле тропомиозина прикреплена молекула другого белка —тропонина (рис.18).Тропонин и тропомиозин являются регуляторными белками и играют важную роль в механизмах взаимодействия актина и миозина при сокращении.