

Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции. В соответствии с этим законом функционируют сложные структуры, например, скелетная мышца. Амплитуда ее сокращений от минимальных (пороговых) величин постепенно увеличивается с увеличением силы раздражителя до субмаксимальных и максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. Поэтому на пороговые раздражители отвечают только те мышечные волокна, которые имеют самую высокую возбудимость, амплитуда мышечного сокращения при этом минимальна. С увеличением силы раздражителя в реакцию вовлекается все большее и большее количество мышечных волокон и амплитуда сокращения мышцы все время увеличивается. Когда в реакцию вовлечены все мышечные волокна, составляющие данную мышцу, дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения.
Закон раздраженияДюбуа-Реймона (аккомодации): раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени. При действии медленно нарастающего раздражителя возбуждение не возникает, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило названиеаккомодации.Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает. Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого - окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще. Способность к аккомодации различных структур неодинакова. Наиболее .высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
Закон силы-длительности: раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать для возникновения возбуждения.
Исследования зависимости силы-длительности показали, что последняя имеет гиперболический характер (рис. 3). Из этого следует, что ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Причиной такой' зависимости является мембранная емкость. Очень "короткие" токи просто не успевают разрядить эту емкость до критического уровня деполяризации. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого действует ток, равный реобазе, и вызывает возбуждение, называетсяполезным временем.
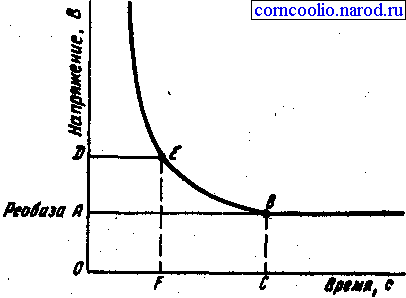
Рис.3. Графическое выражение закона силы-длительности.
В связи с тем, что определение этого времени затруднено, было введено понятие хронаксия - минимальное время, в течение которого ток, равный двум реобазам, должен действовать на ткань, чтобы вызвать ответную реакцию. Определение хронаксии -хронаксимет-рия - находит применение в клинике. Электрический ток, приложенный к мышце, проходит через как мышечные, так и нервные волокна и их окончания, находящиеся в этой мышце. Так как хронаксия нервных волокон значительно меньше хронаксии мышечных волокон, то при исследовании хронаксии мышцы практически получают хронаксию нервных волокон. Если нерв поврежден или произошла гибель соответствующих мотонейронов спинного мозга (это имеет место при полимиелите и некоторых других заболеваниях), то происходит перерождение нервных волокон и тогда определяется хронаксия уже мышечных волокон, которая имеет большую величину, чем нервных волокон.
Закон полярного действия постоянного тока: при замыкании тока возбуждение возникает под катодом, а при размыкании - под анодом. Прохождение постоянного электрического тока через нервное или мышечное волокно вызывает изменение мембранного потенциала покоя. Так, в области приложения к возбудимой ткани катода положительный потенциал на наружной стороне мембраны уменьшается, возникает деполяризация, которая быстро достигает критического уровня и вызывает возбуждение. В области же приложения анода положительный потенциал на наружной стороне мембраны возрастает, происходит гиперполяризация мембраны и возбуждение не возникает. Но при этом под анодом критический уровень деполяризации смещается к уровню потенциала покоя. Поэтому при размыкании цепи тока гиперполяризация на мембране исчезает и потенциал покоя, возвращаясь к исходной величине, достигает смещенного критического уровнями возникает возбуждение.
Закон физиологического электротона: действие постоянного тока на ткань сопровождается изменением ее возбудимости. При прохождении постоянного тока через нерв или мышцу порог раздражения под катодом и соседних с ним участках понижается вследствие деполяризации мембраны - возбудимость повышается. В области приложения анода происходит повышение порога раздражения, т. е. снижение возбудимости вследствие гиперполяризации мембраны. Эти изменения возбудимости под катодом и анодом получили названиеэлектротона(электротоническое изменение возбудимости). Повышение возбудимости под катодом называетсякатэлектротоном,а снижение возбудимости под анодом -анэлектротоном.
При дальнейшем действии постоянного тока первоначальное повышение возбудимости под катодом сменяется ее понижением, развивается так называемая катодическая депрессия.Первоначальное же снижение возбудимости под анодом сменяется ее повышением -анодная экзальтация.При этом в области приложения катода происходит инактивация натриевых каналов, а в области действия анода происходит снижение калиевой проницаемости и ослабление исходной инактивации натриевой проницаемости.
Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации(т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности было названо Н. Е. Введенскимпарабиозом.В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы: уравнительную, парадоксальную, тормозную.
В уравнительную фазупроисходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители - больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100) , то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) - происходит уравнивание ответной реакции.
В парадоксальную-фазупроисходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на частые.
В тормозную фазулабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние.
Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ также связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности.
Механизм развития парабиотического состояния сводится к следующему. При воздействии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость в ответ на раздражение. В участке альтерации инак-тивация натриевых каналов, вызванная повреждающим агентом, суммируется с инактивацией, вызываемой нервным импульсом, и возбудимость снижается настолько, что проведение следующего импульса блокируется.
Введение
ФИЗИОЛОГИЯ - наука о жизнедеятельности организма как целого, так и его частей: тканей, органов Различают: общую, частную прикладную физиологию
Делиться на нормальную и патологическую физиологию
Нормальная физиология изучает деятельность структур и всего живого организма, приспособленности его к внешней среде
Физиология возбудимых тканей
Раздражители - живой клетки реагирует на действие раздр. изменения структур мембраны и протоионозотического свойством клетки. раздражимость - способность живой ткани отвечать раздражитель Раздражимость обладают нервная, мышечная и секреторная ткани их объединяют в понятиеВозбудимость.Возбуждение - это сложный биологический процесс, который характеризуется специфическим изменением процессов обмена веществ, теплообразования, временной деполяризацией мембраны клеток и проявляющийся специализированной реакцией ткани (сокращение мышцы, отделение секрета железой и т. д.).
Классификация раздражителей.:
физические(механические, температурные, звуковые, световые, электрические);
химические(щелочи, кислоты, гормоны, продукты обмена веществ и др.);
информационные
порог раздражения - минимальная сила раздражителя, которая способна вызвать возбуждение.
пороговые- миним.сила способная вызвать ответную реакцию
подпороговыми- если структура не отвечает на раздражение
более сильные - сверхпороговыми
По степени приспособленности биологических структур к их восприятию раздражители делятся на адекватные и неадекватные.
Дистактные
контактные
Первые попытки последовательной разработки учения о "животном электричестве" связаны с именем Л. Гальвани. Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенного на медном крючке, при прикосновении лапок к железным перилам балкона. На основании этих наблюдений Л. Гальвани пришел к выводу, что сокращение мышц лапок вызвано "животным электричеством", которое возникает в спинном мозге и передается по металлическим проводникам к мышцам лапки. Этот опыт в настоящее время известен как первый опыт Гальвани.
Функциональная активная структура - мембрана.
Функции – пограничная (цитоплазма от внеклеточной жидкости)
Биотрансформирующая (барьер для веществ идущих в клетку)
Рецепторная ( мембрана имеет набор структур участвующих в специальных определений физических и химических факторов)
Мембрана образует межклеточные контакты
Транспортная ( переносит разм в-ва в связи с изменение проницаемости)
В состав мембраны двойной слой фосфолипидов, в которой встроены белки и поверхность покрыта мукополисахаридами. Согласно жидкостно-мозаичной модели мембраны молекулы белка могут перемещаться в билипидном слое.
белки делятся на 5 классов:
насосы- расход метаб.энергии АТФ для перемещения ионов и молекул против концетракционных и электрохимических градиентов. Поддерживает необходимый концентрат ионов в клетке
каналы – сам. Транспортный механизм необходимый для переноса веществ через мембрану. Состоит из поры, воротного механизм, сенсора или индикатора и селективного фильтра. Пора это динамическое образование, которое может находиться в открытом или закрытом состоянии. Она образована белком с высокой каталитической активностью. Белок способен переноситься со скоростью в 200 раз больше чем скорость диффузии. Воротный механизм – находится на внутренней поверхности мембраны, состоит из отрицательно заряженных белков способные к конформации (изменение формы). Этот механизм изменяет величину потока ионов. Селективный фильтр на наружной поверхности мембраны , состоит из атомов О2, отрицательно заряженных притягив катионы, отталкиваются анионы. Формируется однонаправленное направление ионов. Сенсор – состоит из положительно заряденых белков.Классификация каналов: селективные – потенциалозависимыми., хемозависимые (рецепторуправляемые), по виду ионов: натриевые, калиевые, кальциевые, хлорные , по скорости: быстрые и медленные.
Неселективные (каналы утечки)
рецепторы
ферменты
структурные (пластические)
мембрана любой живой клетки проницаема для ионов оснований калия, кальция, натрия, и хлора, с помощью активных (идет против электрохимического градиента с затратой энергии и обязательным участием белков переносчиков) и пассивных ( диффузия , осмос, фильтрация; идентичен по концентрации градиенту без затраты энергии из большее в меньшее) транспортов
Состояние живой клетки: покой и возбуждение
Покой характеризуется Поляризация мембраны, т.е. наличие противоположных зарядов по обе стороны. В состоянии покоя мембрана в 10 раз более проницаема для ионов кальция, чем для ионов калия и хлора.
и ионы калия каналах по концентрационному градиенту выходит из клетки потому что в цитоплазме их в 50 раз больше, чем в внеклеточной жидкости. Это вызывает повышение электроотрицательные ионы расположения в цитоплазме. Белки цитоплазмы остаются на поверхности так как не могут пройти в цитоплазму. Клетка омывается внеклеточной жидкостью. Разница растворов ( цитоплазмы и внеклет) только в концентрации ионов. Растворы стремятся уровнять. Ионы калия выходят ионы натрия пытаются попасть каналы пропускают только мелкие ионы (белковые остатки, ионы на крупные) при помощи микроэлектор методики можно измерить величину заряда (мембр. Потенциал) за основу заряда внутренней поверхности мембранные это заряд называется мембранного потенциала покоя. Для разных тканей от 50 до 90 мВ ( со знаком «- «). Величина МПП : заряды, идущие через мембрану ионов и величиной потока ионов, направляющих поток ионов через ионный канал
Кроме того, концентрационного градиента имеется еще и сила, обеспечивающих перемещение ионов от одноименного к противоположному электрическому градиенту. А механизм, получил название натрий-калиевого насоса.Он обеспечивает активный транспорт ионов натрия во внеклеточную жидкость и введении в цитоплазму ионов калия. Переход из покоя в возбуждение характеризируется -деполяризациямембраны и выражается в виде генерации клеточного потенциала. В зависимости от силы раздражения потенциал может быть местным –локальный ответ. Такой потенциал не распространяется и затухает вблизи распространения. Возникаетдеполяризациисвязи с повышением проницаемости для ионов натрия идущих в клетку. Однако во время локального ответа мембранный потенциал не достигает критического уровня т.е. момента когда не распространяется возбуждение переходит в распространяющееся.
КУДне распространяющейся переход в распространяющееся . и возбуждение переходит на близ лежащие участки. Расстояние между мембранного потенциала и критического уровня называется пороговым популяциями. И возбуждение обратно существует . При действии на ткань раздражение пороговой силы повышает проницаемость для ионов натрия, т.к. открываются потенциально зависимые натрий- каналы и ионы натрия поступают внутрь клетки. Потенциально зависимые натрий- каналы имеют два типа ворот : М – активационными, и Н инактивационными
В состоянии покоя активация ворот закрыты, а инактивационные открыты. После нанесения раздражения активация ворот открыты и натрий устремляется внутрь клетки. К- каналы очень инертны
Постепенно натрий ток возрастает в 500 раз по сравнению состояния покоя и начинается преобладание на выход К-током. Процесс нарастает медленно до критического уровня деполяризации и процесс становится не управляем. Натрий лавиной устремляется в клетку и деполяризует мембрану зарядом «+». Ионный ток проходит «О» отметку, и заряжается «+». Натрий – каналы инактивируют (закрывают) и прекращают пропуск натрия, а калий только набирает и «+» заряду выход наружу. Достигает критического уровня К-каналы инактивируется, процесс замедляется. Насос выкачивает натрий и закачивает калий восстанавливает ионный баланс. Клетка вернулась в состояние покоя необходимо время для снижения силы натрий-калиевого насоса и натрий перекачивает на поверхность и возникает гиперполяризация и восстанавливается ионная ассиметрия.
Нанесение подпорогового раздражения возникает локальный ответ. Он способен к суммации (выход за критический уровень)
ВЫВОД: электрические явления на клетку мембраны . Для понятия реакции ткани на раздражение.
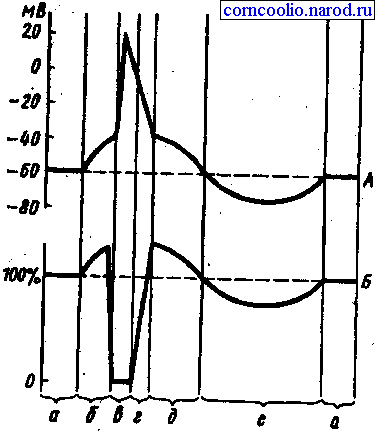
Рис. 2. Соотношение одиночного цикла возбуждения (А) и фаз возбудимости (Б). А: а - мембранный потенциал покоя; б - предспайк, локальный ответ, ВПСП; в - спайк, потенциал действия, деполяризация и инверсия; г - потенциал действия, реполяризация; д - отрицательный следовой потенциал, следовая деполяризация; е - положительный следовой потенциал, следовая гиперполяризация. Б: а - исходный уровень возбудимости; б - фаза первичной экзальтации, повышенная возбудимость; в - фаза абсолютной рефрактерности; г - фаза относительной рефрактернности; д - фаза вторичной экзальтации; е - фаза вторичной рефрактерности
Изменение возбудимости в разные фазы потенциального действия. В разные моменты токи движутся неравномерно. (отщепляясь АТФ выводит энергию и открывает ворота.
1. состояние локального ответа – ворота открыты мало АТФ не расщепилась, энергии очень много, наносим дополнительное раздражение – фаза возбудимости (супернормальная)
2 все каналы открыты, наносим дополнительное раздражение, ответа не будет (фаза абсолютной невозбудимости) фаза абсолютной рефрактернности
3 инактивация каналов локального ответа накапливается на мембране, наносит раздражение если раздражение пороговой силы ответа не будет если сверх пороговый то это фаза относительной рефрактернности
Отрицательно средовой потенциала. – знергия АТФ на мембрану накапливается следовательно на подпороговое раздражение получают максимальный ответ - фаза вторичной экзальтации. Гиперполяризация мембраны наносится раздражение продолжается выход ионов калия; заряд не изменяется ответ слабый (фаза субнормальной возбудимости