

Сегмент 27. Внутриклеточная саморегуляция
В клетке для поддержания гомеостаза используются в основном химические (молекулярные) механизмы регуляции. Наиболее важна регуляция генов, от которых зависит производство белков, в том числе многочисленных и разнообразных ферментов.
Самая простая модель для демонстрации генного гомеостаза - регуляция выработки фермента для расщепления пищевого сахара у кишечной палочки. Эта бактерия является типичным гетеротрофом и поглощает из внешней среды несложные органические вещества, в том числе молочный сахар лактозу. Для расщепления и усвоения лактозы с определенного структурного гена, входящего в состав лактозного оперона (ген вместе с регуляторной областью) синтезируется информационная РНК и, далее, фермент (см. центральную догму молекулярной биологии - сегмент 22, рис. 13). Если сахар в среде отсутствует, фермент не вырабатывается, а при добавлении сахара активируется ген и идет синтез фермента. Но как только весь сахар будет клеткой использован, ген перестает работать. Как клетка узнает о присутствии сахара и его расходовании? Как оберегает свои гены от бесполезной работы и траты энергии? Регуляция генов у бактерий, как у всех прокариот, в целом организована гораздо проще, чем в эукариотных клетках. Оказывается, лактозный оперон у кишечной палочки работает по принципу отрицательной обратной связи, где в роли регуляторного «клапана» выступает особый участок оперона - оператор, а в роли регулятора сам пищевой субстрат - лактоза (рис. 19). Лактоза, поступившая в клетку, сама раскрывает структурный ген, используя для этого в качестве ключика операторный участок. Исчезновение лактозы автоматически приводит к закрытию гена.
Рис. 19
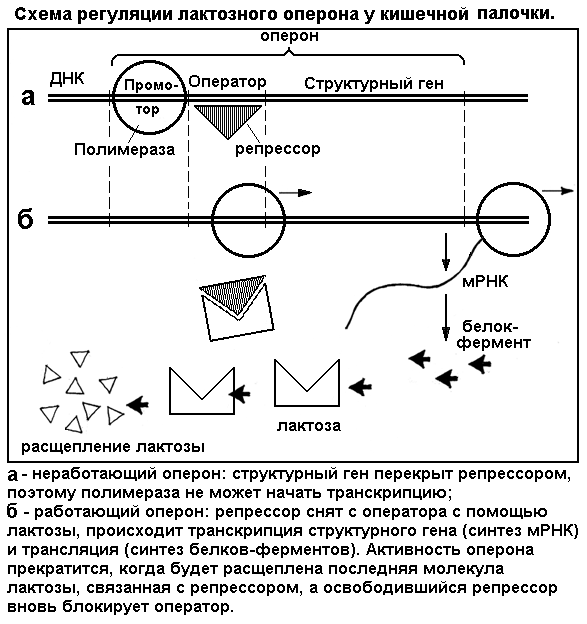
Лактозный оперон- участок молекулы ДНК - состоит из трех частей: промотора, оператора и структурного гена.Промотор- стартовый участок гена, сюда садится фермент РНК-полимераза, ведущий транскрипцию.Оператор - пусковой барьер, в отсутствие лактозы закрытый специальным белком-репрессором.Структурный ген(точнее - здесь находится цепочка, семейство генов) - основной участок ДНК, кодирующий и производящий через иРНК нужный белок-фермент. Пока оператор связан с белком-репрессором, полимераза не может стартовать и структурный ген не работает, синтез фермента отсутствует (см. рис. 19 а). Когда в клетку попадает лактоза, одна ее молекула связывается с репрессором и отнимает его от оператора. Теперь путь полимеразе открыт, идет синтез иРНК (транскрипция) и, далее, синтез соответствующего белка-фермента (трансляция) (рис. 19 б). Ферменты расщепляют поступивший в клетку сахар и в последнюю очередь ту его молекулу, которая связана с репрессором. Но когда будет переварена и эта последняя молекула, белок-репрессор освобождается и вновь блокирует оператор. Производство иРНК и фермента прекращается до поступления новой порции сахара. По своей простоте система регуляции гена концентрацией субстрата похожа на простые технические регуляторы. Напомним, однако, что у эукариот регуляция генной активности более сложная. Она включает возбуждение клеточных рецепторов гормонами или другими биологически активными веществами, запуск каскада реакций вторичных мессенджеров, которые поступают в ядро и избирательно активируют гены.
Другой пример простых саморегулирующихся систем, использующих обратную отрицательную связь, представляют ферментативные цепи, ингибируемые конечным продуктом (рис. 20). Такие цепи обычно локализуются на поверхности внутриклеточных или наружных мембран и проводят комплексную переработку сложного субстрата в простой продукт. Суть регуляции состоит в том, что конечный продукт имеет стереохимическое сродство с первым ферментом. Связываясь с ферментом, продукт ингибирует (подавляет) его активность, так как полностью искажает его третичную структуру. Работает следующий регуляторный цикл. При повышении концентрации конечного продукта выше необходимого уровня его избыток ингибирует ферментную цепь (для этого достаточно остановить самый первый фермент). Ферментация прекращается, а свободный продукт расходуется на нужды клетки. Через некоторое время возникает дефицит продукта, блок с ферментов снимается, цепь активируется, и производство продукта снова растет.
Рис. 20

Третий пример- поддержание внутриклеточного осмотического гомеостаза. В сегменте 19 мы говорили о механизме возникновения нервных импульсов и отмечали важную роль ионов натрия, концентрация которых снаружи клетки должна поддерживаться на более высоком уровне, чем внутри. Благодаря натриевым насосам, встроенным в мембрану клетки, удерживается нужный градиент ионов. Как только клетка получает избыток натрия, активируется натриевый насос (его фермент, расщепляющий АТФ и дающий энергию). Натрий выкачивается, его концентрация в клетке падает, что служит сигналом для отключения насоса.
Аналогично в клетках растений с помощью плазмалеммы (наружной мембраны) и вакуолей регулируется состав солей и питательных веществ. Плазмалемма обеспечивает приток в клетку необходимых ионов и воды из внешней среды и выделение балластных и избыточных ионов водорода, натрия, кальция. Мембрана вакуоли регулирует поступление в протоплазму запасных субстратов из вакуоли при их недостатке и удаление в вакуоль - при избытке.
Во всех рассмотренных случаях действует один и тот же принцип - саморегуляция системы на основе обратной отрицательной связи. Заметим, однако, что регулируемые параметры - концентрация солей, питательных веществ, конечного продукта ферментации или продукта генной активности - не бывают абсолютно постоянными, они поддерживаются в допустимых границах. В каждом случае это свои физиологические границы, позволяющие нормально осуществлять клеточные функции. Аналогичный принцип мы увидим и на организменном уровне.