

Місцеві гормони або активні фактори
На ряду з основною медіацією – адренергічною і холінергічною, завдяки якій здійснюється передача симпатичних і парасимпатичних впливів на ефекторні структури дуги автоматичного рефлексу. Організм має більшу кількість біологічно активних речовин. Ці речовини, які мають назву місцевих гормонів або активних факторів здатні надавати значний вплив на дію і тонус структур автономної нервової системи. Серед них слід назвати простогландини і плазмакініни. В наш час розрізняють 14 різновидів простогландинів, які розділяють на 4 групи:
e, F, а, B.
Вихідним продуктом синтезу простогландинів є ліпоєва кислота. Основним місцем локалізації в клітині є жировий шар мембран і оболонок. В структурах автономної нервової системиміститься вимірювана кількість простогландинів, вони можуть виділятись спонтанно чи під впливом стимуляції. Більш ймовірною роллю простогландинів є координація нейрогуморальної передачі і участь в механізмах, вивільнюючих і виділяючих медіаторів.
Плазмокініни – є поліпептидами, які утворюються із плазматичних глобулінів під впливом протеолітичних ензимів. Вони містяться в тканинах різних органів, плазмі крові, слині. Зараз найбільш відомі 3 біологіно активних плазмокініни – брадікініни, калідін, метіонілкалійдін.
Плазмокінінам приписується активна участь у розвитку патологічних станів, таких як мігрень, англоневротичний набряк, панкреатит, травматичний шок. Особливе місце відноситься у генезі запальної реакції, так як у результаті місцевого вивільнення плазмокінінів виникає гіперемія, внаслідок підвищення проникливості судинної стіінки – набряк, внаслідок подразнення респіраторного апарату - біль. Усе це є основою для розгляду плазмокінінів у якості комплексного медіатора запальної реакції.
Вищі центри регулюючі дію автономної системи
В нервовій системі високоорганізованих організмів на ряду з вузькою спеціалізацією окремих клітин існує спеціалізація більш загального характеру – спеціалізації 2 крупних відділів. Функцією першого цереброспінального є встановлення необхідних зв’язків і відношення до отчуючого зовнішнього світу. Функцією другого – автономного – забезпечення правильного протікання процесів життєдіяльності всередині організму. Діяльність цих відділів тісно пов’язана одна з другою і взаємно одна одну доповнюють. Для корекції діяльності існує ще вища ступінь централізації – апарат координації. Таким апаратом є підкіркові центри та кора головного мозку. Надсегментарні апарати особливістю яких є відсутністю специфічних морфо-фукціональних рис притаманних периферичному відділу автономної нервової системи. Розташовані в основному на лімбікоретикулярному рівні. У склад лімбічної системи входять наступні філогенетично розрізняємі структури:
підкіркові утворення – мигдалевидний комплекс, прозора перетинка, передні ядра зорового бугра і гіпоталамуса, ретикулярна фармація середнього мозку.
Кора великих півкуль – лімбічна ( внутрішня доля), складається з 2 звивин – сводових розташованих над мозолистим тілом(врехня лімбічна доля) і гіпокамповий вистилаючий нижню внутрішню поверхню кори( нижній лімбічний відділ). Головним підкорковим цетром інтеграції автономної функції вважається гіпоталамус: тут розташовуються структури регулюючі основні життєві функції. Центри гіпоталамуса не є не симпатичними, не парасимпатичними, вони об’єднують регуляції функцій обох частин автономної нервової системи в залежності від рівня приходящої до них аферентації. Гіпоталамус має виключне значення в нейрогуморальній і гормональній регуляції. До сих пір гіпоталамус традиційно розглядається як вищий центр регуляції вісцеральних функцій.
Для підтримання гомеостазу і виконання своїх складних функцій автономної системи на ряду з нормальними нейронами має у своєму складі особливі клітини, в яких прийом інформації здійснюється звичайним шляхом, а відповіді ендокринним способом, такі клітини мають назву – трансдуктів. До них відносяться:
α-хромафінові клітини мозкового шару наднирників, які відповідають на холінергічний передаччик симпатичного волокна виділенням адреналіну і норадреналіну;
юкстагломерумерні клітини нирок, які на адренергічний передаччик симпатичного волокна відповідає виділенням в кровоносне русло реніну;
нейрони супраоптичного і паравентрикулярного ядер гіпоталамуса, реалізуючі на холінергічний і адренергічний та інші медіатори, віиділенням за допомогою нейрогіпофізу вазопресина і оксітоцина;
нейрони ядер гіпоталамуса, виділяють в судинну систему фактори регуляції
Тема 2. Морфофункціональні особливості симпатичної та парасимпатичної інервації різних органів. Еферентна інервація серця, гладенько-м’язових клітин травного тракту, кровоносних судин, секреторних та інкреторних залоз.
Симпатична і парасимпатична нервові системи відрізняються за розміщенням гангліїв. Ганглії симпатичної нервової системи, залежно від їх локалізації, поділяють на вертебральні і превертебральні. Вертебральні симпатичні ганглії розміщені по обидва боки хребта, утворюючи симпатичні нервові ланцюжки, або пограничні стовбури. У цих гангліях переривається більша частина симпатичних нервових волокон. Постгангліонарними нейронами цих гангліїв здійснюється симпатична іннервація голови (очей, слинних залоз і судин), органів грудної порожнини (бронхів, серця) і шкіри (судин і потових залоз).
Менша частина симпатичних нервових волокон переривається не у пограничних стовбурах, а у превертебральних гангліях. Вони більш віддалені від хребта, але і знаходяться на деякій відстані від ефекторних органів. До них належать сонячне сплетення, верхній і нижній брижові вузли, що знаходяться у черевній порожнині. Постгангліонарні нейрони цих гангліїв іннервують шлунок, тонкий і товстий кишечник, печінку, сечовий міхур і статеві органи.
Ганглії парасимпатичної нервової системи розміщені у внутрішніх органах (або поблизу них), тому називаються внутрішньоорганними. Вони наявні у серці, бронхах, середній і нижній частині стравоходу, шлунку, кишечнику, жовчному і сечовому міхурах, екзокринних і ендокринних залозах. Аксони прегангліонарних парасимпатичних нервових волокон довгі і доходять до ефекторних органів не перериваючись. Головним джерелом парасимпатичних впливів служать волокна блукаючого нерва, які іннервують органи грудної і черевної порожнин.
Більшість внутрішніх органів отримує подвійну рухову іннервацію - симпатичну і парасимпатичну. Однак симпатична іннервація більш поширена, ніж парасимпатична. Симпатичну рухову іннервацію отримують фактично всі органи і тканини. Парасимпатичну рухову іннервацію не отримують скелетна мускулатура, більша частина кровоносних судин і матка.
Вегетативні ганглії відіграють важливу роль у розподілі нервових впливів, які реалізуються через них. Кількість нейронів у вегетативних гангліях більша, ніж прегангліонарних нервових волокон. Отже, прегангліонарне нервове волокно утворює синапси з багатьма нейронами гангліїв. За рахунок цього досягається розширення зони впливу прегангліонарних волокон.
Вегетативні ганглії - це не тільки передавальні структури, а й винесені на периферію рефлекторні центри. Експериментально доведено наявність місцевих
периферичних рефлексів, які здійснюються інтрагуморальними вегетативними гангліями. За участю таких рефлексів регулюється діяльність серця, шлунково-кишкового тракту.
Багато внутрішніх органів після симпатичної і парасимпатичної денервації продовжують виконувати свої функції. Це означає, що вони мають власний механізм нервової регуляції. Раніше третій відділ автономної нервової системи називали ентеральним. Рефлекторні дуги ентеральної нервової системи замикаються у підслизовому та міжм'язовому нервових сплетеннях кишок. Сьогодні наявність третього відділу автономної нервової системи передбачають і в інших порожнистих внутрішніх органах, називаючи його метасимпатичним.
Метасимпатична нервова система містить різні типи нейронів, різні медіатори! системи, через неї симпатична і парасимпатична нервові системи модулюють активність внутрішніх органів. Метасимпатична нервова система найменш залежна від центральної нервової системи і служить базовим рівнем іннервації (А.Д.Ноздрачев, 1991). Багато центрів вегетативної нервової системи перебувають постійно у стані активності (тонус центрів). Наприклад, після двобічної ваготомії серцева діяльність прискорюється, внаслідок звільнення від постійного гальмівного впливу центрів блукаючих нервів.
Чутливі ланки рефлекторних дуг вегетативної нервової системи починаються рецепторами вісцеральних органів (інтерорецепторами), які сприймають механічні та хімічні стимули. Інтерорецептори розміщуються у стінках практично всіх внутрішніх органів {серце, судини, легені, нирки, сечовий міхур, печінка й інші).
Механорецептори внутрішніх органів часто називають барорецепторами. Насправді вони є типовими механорецепторами, оскільки їхнє збудження пов'язане не з тиском, а з розтяганням стінки судин або кишечника. Хеморецептори збуджуються в основному під час змін рН середовища. У внутрішніх органах наявні і больові рецептори.
Вісцеральну сенсорну інформацію проводять у нервові центри спинного і довгастого мозку волокна груп А, В і C. У дугах автономних рефлексів перемикання аферентних сигналів на рухові нейрони може здійснюватись і у вегетативних гангліях.
Симпатична і парасимпатична нервові системи спричиняють у більшості органів протилежні ефекти, що свідчить про антагоністичний характер їхнього впливу. Наприклад, симпатична нервова система підсилює серцеву діяльність, її парасимпатична - гальмує. Наявні і винятки з цієї закономірності. Наприклад. слиновиділення стимулює і симпатична, і парасимпатична нервові системи. Крім того, наявні органи, які отримують тільки симпатичну або тільки парасимпатичну рухову іннервацію (табл. 2).
Таблиця 2. Вплив вегетативної нервової системи на діяльність внутрішніх органів
|
00Орган |
Симпатична нервова |
Парасимпатична нервова | ||
|
|
система |
система | ||
|
1 |
2 |
3 | ||
|
Око |
|
| ||
|
Зіниця |
Розширення |
Звуження | ||
|
Ціліарні м'язи |
Не проявляється |
Скорочення | ||
|
Слізні залози |
Не проявляється |
Секреція | ||
|
Слинні залози |
Секреція в'язкої слини |
Секреція водянистої слини | ||
|
Потові залози |
Потовиділення (медіатор ацетилхолін) |
Не проявляється | ||
|
Бронхи |
Розширення |
Звуження | ||
|
Міокард |
|
| ||
|
Ритм скорочень |
Прискорення |
Сповільнення | ||
|
Сила скорочень |
Збільшення |
Послаблення | ||
|
Кровоносні судини |
|
| ||
|
Коронарні |
Розширення |
Звуження | ||
|
Мозку |
Розширення |
Звуження | ||
|
Шкіри |
Звуження |
Не проявляється | ||
|
Скелетних м'язів |
Розширення |
Не проявляється | ||
|
Черевної порож- |
Звуження |
Не проявляється | ||
|
нини |
|
і | ||
|
Артеріальний тиск |
Підвищення |
Зниження | ||
|
Шлунок |
|
| ||
|
Секреція |
Гальмування |
Підсилення | ||
|
Кишечник |
|
| ||
|
Тонус і моторика |
Гальмування |
Підсилення | ||
|
Сфінктери |
Скорочення |
Розслаблення | ||
|
Секреція |
Гальмування |
Збудження | ||
|
Підшлункова залоза |
Не проявляється |
Збудження | ||
|
Печінка |
Глікогеноліз Глюконеогенез |
Не проявляється : : | ||
|
Жовчні шляхи |
Розслаблення |
Скорочення | ||
|
Сечоводи |
Гальмування скорочень |
Підсилення скорочень | ||
|
Сечовий міхур |
■ |
| ||
|
М'яз, який вишто- |
Розслаблення |
Скорочення | ||
|
вхує сечу |
|
| ||
|
Сфінктер |
Скорочення |
Розслаблення | ||
|
Статеві органи |
Еякуляція |
Ерекція | ||
|
|
|
| ||
|
|
|
| ||
|
1 |
2 |
3 | ||
|
М'язи, які піднімають |
Підсилення скорочень |
Не проявляється | ||
|
волосся |
|
| ||
|
Основний обмін |
Підвищення |
Не проявляється | ||
|
Зсідання крові |
Підвищення |
Не проявляється | ||
|
Психічна активність |
Підвищення |
Не проявляється | ||
Якщо якому-небудь внутрішньому органу притаманна автоматія (серце, шлунково-кишковий тракт), симпатична або парасимпатична нервова система підсилює або послаблює його діяльність. У цьому випадку говорять про коригуючий вплив вегетативної нервової системи. Якщо ж орган не працює автоматично, а збуджується імпульсами симпатичної або парасимпатичної нервової системи, говорять про пусковий вплив вегетативної нервової системи.
Оскільки симпатичні постгангліонарні нейрони характеризуються більш широкою зоною іннервації, їхнє збудження приводить до генералізованих реакцій. Сумарний ефект симпатичної нервової системи полягає у гальмуванні активності багатьох внутрішніх органів і стимуляції серця і скелетних м'язів, тобто у підготовці організму до поведінки типу боротьби або втечі. Загальне збудження симпатичної нервової системи допомагає організму справитись із стресовими ситуаціями. Симпатична нервова система активує діяльність нервової системи в цілому, активує захисні сили організму (імунні процеси, бар'єрні механізми, зсідання крові, процеси терморегуляції) і мобілізує енергетичні та інтелектуальні ресурси організму, коли виникає загроза його існуванню.
Парасимпатичні постгангліонарні нейрони знаходяться у самих органах, володіючи обмеженими зонами іннервації. Тому вважають, що парасимпатичні нейрони чинять місцеву дію на функцію внутрішніх органів. Загалом парасимпатична нервова система відповідає за відновлення ресурсів і підтримання гомеостазу - зниження частоти серцевих скорочень, артеріального тиску і стимуляцію функцій травної системи. Парасимпатична нервова система діє або безпосередньо на органи і тканини (кільцевий м'яз райдужної оболонки, слинні залози), або через метасимпатичну нервову систему.
Отже, симпатичну нервову систему розглядають як систему тривоги, мобілізації захисних сил і ресурсів для активної взаємодії з факторами середовища, що дестабілізують сталість внутрішнього середовища. Пара- і метасимпатична нервові системи забезпечують "відновлення і підтримання сталості внутрішнього середовища.
Тема 3. Фізіологічні реакції внутрішніх органів та систем органів при збуджені симпатичних і парасимпатичних нервів. Уявлення про антагоністичні та синенергічні відносини між симпатичною та парасимпатичною системами.
По уявленням французького фізіолога М. Біла (початок ХІХ століття) функції тваринного організму поділяли на дві групи: тваринні (анімальні, соматичні) і вегетативні (рослинні). Перші включали рухові реакції скелетних м’язів і сприйняття зовнішніх подразників, другі – решту функцій тіла – живлення, ріст, розмноження і т.п. У відповідності з цим було встановлено фізіологічний принцип поділення нервової системи на соматичну і вегетативну (від лат. Vegetativus – рослинний).
Таким чином, термін «вегетативна нервова система” існує майже два віки, віки, а її виділення безумовно стимулювало і клінічні, і фізіологічні, і біохімічні підходи до вивчення особливостей і закономірностей функціонування як соматичної, так і вегетативної систем. Треба зазначити, що термінологія „вегетативна нервова система” влаштовувала не всіх вчених. В зв’язку з чим були початі багато чисельні спроби до її покращення.
Французький фізіолог Клод Бернар в основу виділення вегетативної нервової системи поклав нову ознаку – не довільну іннервацію. Ознака не довільності дозволила англійському фізіологу В. Гаскеллу звернути увагу на наявність в організмі двох типів м’язової тканини, які підпорядковуються відповідно “довільній” і “не довільній” іннервації. В межах не довільної іннервації виявилась іннервація м’язів судин, шкірних утворень, внутрішніх органів. Ознака не довільності не є єдиною, при віднесенні нервового утворення до того чи іншого відділу. Гаскелл показав також існування хімічної чутливості м’язів, що іннервуються не довільно нервовою системою, частина з яких реагує скороченням на застосування адреналіну. Це дозволило йому розділити не довільну нервову систему на власне симпатичну (адреналову) і нервову систему внутрішніх органів, виділив при цьому краніальний, тораколюмбальний і сакральний відділи. Не довільну нервову систему він запропонував позначити як „вісцеральна система”. Безперечно, що вегетативно-вісцеральна регуляція дуже важлива, але вегетативна іннервація захоплює всі тканини організму і не являється лише „вісцеральною”.
Співвітчизник Гаскелла Дж. Н. Ленглі встановив різницю в будові довільної і не довільної іннервації. Він показав, що довільна соматична іннервація здійснюється однонейронним шляхом – тіло нервової клітини лежить в ЦНС, а її аксон досягає виконавчого органу (скелетного м’яза). В той же час шлях не довільної вегетативної іннервації представлено двома нейронами, перший з яких знаходиться в ЦНС, а другий – в периферійному ганглії.
Цю не довільну частину нервової системи Ленглі назвав „автономною нервовою системою”, підкреслив при цьому її велику ступінь незалежності від ЦНС. Він рахував цю систему як цілком еферентну. В автономній нервовій системі Ленглі виділив симпатичну нервову систему (тораколюмбальний відділ) та позначив решту утворень (краніальний і сакральний відділи) терміном парасимпатична. При цьому нервові сплетіння стінки кишки він окремо назвав ентеральною системою.
Термін „автономна нервова система” обмежено у використанні, так як вегетативна автономія є відносною, а число публікацій про довільне управління, можливостях довільного регулювання постійно зростає (практика йогів, аутогенне тренування, використання біологічного зворотного зв’язку). Були пропозиції використати виходячи з анатомічних даних термін „вузлова нервова система”. В результаті життєвими виявились два терміни: „вегетативна нервова система”, що використовується на території СРСР і німецько-, франкомовних державах, і „автономна нервова система”, що розповсюджується в англомовних державах. У зв’язку з тим, що англійська мова все більш домінує, другий термін „автономна нервова система” починає домінувати. Чітко уявляємо собі, що обидва визначення є синонімами, ми традиційно застосовували поняття „вегетативна нервова система”.
Особливу роль вегетативної нервової системи в організмі обґрунтував Л.А. Орбелі. Дані його школи показали, що симпатична іннервація впливає на функціональний стан всіх без виключення органів і тканин. В тому числі і відділів ЦНС. Таким чином було сформульовано основний принцип її функціонування в організмі – адаптаційно-трофічний характер її впливів.
Головна функція вегетативної нервової системи полягає в підтримці сталості внутрішнього середовища (гомеостазу) при різноманітних впливах на організм. В той же час ця система реагує також на діяльність органів і систем, які безпосередньо не приймають участь в підготовці гомеостазу (наприклад, статевих органів і внутрішніх м’язів ока).
Вегетативна (автономна) нервова система включає морфологічно і функціонально різні відділи: симпатичний, парасимпатичний і метасимпатичний. Регуляція вісцеральних функцій здійснюється вегетативною нервовою системою за допомогою рефлексів, що одержали назву вегетативних. Структурною основою вегетативних рефлексів, як і соматичних, є рефлекторна дуга.
Нервові центри вегетативних рефлексів симпатичної і парасимпатичної систем розташовані в різних відділах центральної нервової системи. Так, нервові центри симпатичної нервової системи мають торако-люмбальну локалізацію у спинному мозку, нейрони цих центрів розташовані в бічних рогах сірої речовини від верхнього грудного до 4 - 5 поперекового сегментів (T1-L5). Центри парасимпатичного відділу мають краніо-сакральну локалізацію. Краніальні центри знаходяться в ядрах III, VII, IX, Х пар черепно-мозкових нервів, а сакральні — у бічних рогах трьох сегментів крижового відділу спинного мозку (рис. 1.1).
Особливістю вегетативної нервової системи є локалізація еферентного нейрону нервового центра, винесеного за межі центральної нервової системи і розташованого у вегетативних гангліях. У нервових центрах соматичних рефлексів і вставні, і еферентні нейрони розташовані в межах мозку, у вегетативних центрах вставні нейрони локалізовані в мозку, а еферентні — у гангліях. Для симпатичного відділу це превертебральні ганглії і симпатичні стовбури, що лежать по дві сторони хребетного стовбуру, для парасимпатичного відділу — паравертебральні ганглії і нервові вузли, що лежать поблизу органів, які вони іннервують. Нервові провідники, що відходять від вставних нейронів мозку і виходять у напрямку до гангліїв, одержали назву прегангліонарних, вони утворюють синапси на еферентних нейронах гангліїв, тому їх синапси називають прегангліонарними.
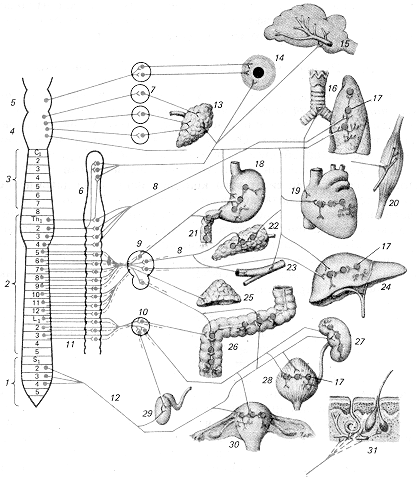
Рис. 1.1. Вегетативна нервова система (загальний план будови еферентної частини): 1 — крижовий відділ спинного мозку (S), 2 — торако-люмбальний відділ спинного мозку (Тh-L), 3 — шийний відділ спинного мозку (З), 4 — довгастий мозок, 5 — середній мозок, 6 — прикордонний симпатичний стовбур (паравертебральні ганглії), 7 — парасимпатичні вузли голови, 8 — постгангліонарні волокна, 9 — сонячне сплетіння і його вузли, 10 — каудальне брижове сплетення і його вузли, 11 — прегангліонарні волокна, 12 — тазовий нерв, 13 — слинні залози, 14 — внутрішні м'язи очей, 15 — судини головного мозку і його оболонок, 16 — трахея, бронхи, легеня, 17 — функціональні модулі метасимпатичної нервової системи, 18 — шлунок, 19 — серце, 20 — скелетний м'яз, 11 — дванадцятипала кишка, 22 — підшлункова залоза, 23 — судини внутрішніх органів, 24 — печінка, 25 — надниркова залоза, 26 — товста кишка, 27 — нирка, 28 — сечовий міхур, 29 — внутрішня частина чоловічих статевих органів. 30 — матка, 31 — шкіра
Нервові провідники, що відходять від нейронів та гангліїв еферентно, називають постгангліонарними, вони утворюють синапси на клітинах органів, або ефекторах які вони регулюють. Ці синапси називають постгангліонарними. Тому в симпатичному відділі прегангліонарні нервові волокна, як правило, короткі, а постгангліонарні — довгі. У парасимпатичному відділі, навпаки, прегангліонарні волокна, як правило, довгі, а постгангліонарні — короткі.
Рецептори вегетативних рефлексів, які розташовані у внутрішніх органах, стінках кровоносних і лімфатичних судин, шкірі і м'язах називаються інтероцепторами. Усі вони відносяться до первинно чутливих рецепторів, тобто є кінцевими утвореннями аферентних нервових волокон.
Аферентні волокна проходять до нервових центрів або в складі вегетативних нервів, що містять як аферентні, так і еферентні провідники (наприклад, блукаючий нерв), або в складі соматичних аферентних нервів від м'язів та шкіри, тому такі нерви називають ще змішаними. У центральній нервовій системі аферентні провідники утворюють велику кількість синапсів на вставних нейронах, при цьому за рахунок значної дивергенції інформація, що надходить, переключається не тільки по напрямку еферентних нейронів – гангліїв, але і на висхідні шляхи до вищих вегетативних центрів головного мозку, кори великих півкуль та на нейрони соматичних рефлекторних дуг. Це забезпечує інтеграцію вегетативних рефлексів між собою і соматичними рефлексами для забезпечення соматичних функцій.
Прегангліонарні провідники симпатичної нервової системи залишають спинний мозок у складі передніх корінців сегментів спинного мозку і через білі сполучні гілки входять у превертебральні ганглії та симпатичні або прикордонні стовбури, де утворюють прегангліонарні синапси на нейронах гангліїв. Прегангліонарні провідники парасимпатичної нервової системи виходять з центрів краніального відділу в складі черепно-мозкових нервів.
Постгангліонарні еферентні провідники симпатичного відділу виходячи з гангліїв, або входять у соматичні нерви через сірі сполучні гілки й у їхньому складі проходять до ефекторів, або утворюють самостійні симпатичні нерви. Короткі постгангліонарні парасимпатичні волокна гілкуються в товщі самого органу де утворюють синапси.
Еферентні симпатичні волокна іннервують практично усі без винятку тканини й органи, тоді як парасимпатичні волокна не іннервують скелетні м'язи, матку, головний мозок, кровоносні судини шкіри, черевної порожнини і м'язів, органи почуттів і мозкову речовину надниркових залоз.
Еволюція вегетативної нервової системи починається, певно, з безхребетних. У деяких безхребетних (аннелід та ін.) від соматичної нервової системи відокремлюються нервові елементи, пов’язані з кишковою трубкою. Вони утворюються з клітин глоткових вузлів і формують самостійні ганглії. У членистоногих існує вже достатньо чітко диференційована вегетативна нервова система з вузлами і нервовими стовбурами, що йдуть до м’язів шлунка. У бабки, наприклад, виділяють краніальний, тулубовий та каудальний відділи. Тулубовий відповідає симпатичній, краніальний і каудальний – парасимпатичній нервовій системі хребетних. У нижчих черепних тварин сформованих вегетативних гангліїв не виявлено, хоча існують вісцеральні гілки спинномозкових нервів.
У низькоорганізованих хребетних, таких як міноги, акули, скати по ходу симпатичних сплетінь, що іннервують травний канал і починаються від вісцеральних гілок спинномозкових нервів, виникають гангліозні скупчення клітин. Вони розташовуються дорзальніше аорти відповідно кожній парі спинномозкових нервів. Від гангліїв відходять гілки до внутрішніх органів і серця. В товщі органів ці гілки формують сплетіння з гангліозними клітинами. Таким чином, на цьому етапі еволюції виникнення частина вегетативної нервової системи – матасимпатична.
В ряду кісткових риб утворюється парний пограничний симпатичний стовбур зі зв’язками, що характерні для вищих хребетних, краніально досягає хвоста. В подальшому в ході еволюції будова вегетативної нервової системи ускладнюється, відрізняючись розмаїтістю у різних груп. Симпатичний стовбур рептилій, наприклад, включає до 27 пар гангліїв з багато членистими зв’язками, інтрамуральні сплетіння внутрішніх органів добре диференційовані, хоча кількість клітин у вузлах ще незначна. У птахів прегангліопарні волокна залишають спинний мозок у складі вентральних корінців.
По мірі філогенетичного розвитку ускладнюється і клітинній склад гангліїв. Якщо у низькоорганізованих хребетних тварин ще нема чітких специфічних різниць між нервовими клітинами, то у більш високоорганізованих представників класу риб та особливо у ссавців є вже три їх типа: рецептори, еферентні і асоціативні.
Згідно А. А. Заварзіна, у процесі розвитку організму з появою білатеральної симетрії і диференціації головного та хвостового кінців почався поділ нервової системи на соматичну й автономну. Автономну нервову систему підрозділяють на симпатичний, парасимпатичний і метасимпатичний відділи. З цієї системи виділилася частина основного тричленного апарату, що зберегла свій зв'язок із соматичною системою. У зв'язку з більш одноманітною і менш складною організацією вісцеральних органів, чим органи руху, з'явилися особливі властивості, які розвивалися із загальних властивостей єдиної нервової системи. Таким шляхом виникла симпатична, парасимпатична і метасимпатична нервова система.
Ембріональним джерелом клітин вегетативної нервової системи у ссавців служить гангліозна пластинка, що підрозділяється на соміти, які дають згодом симпатичну і парасимпатичну нервову систему. Їхня периферична частина, а також метасимпатична нервова система утворилися в результаті подальшої міграції нейробластів у стінки внутрішніх органів.