

Protistologia
.pdf48 Эволюция и таксономия
отной и прокариотной клеткой. Общая схема этого процесса описана в уже упо мянутой теории стадийного эндосимбиоза, которая утверждает, что возникно вение митохондрий от такой симбиотической ассоциации было реализовано только однажды. Вопрос о том, возник ли ли гидрогеносомы (которые встреча ются только у амитохондриальных про тистов) в результате преобразования митохондрий или от эндосимбионтов, давших начало и митохондриям, и гидрогеносомам, пока не может быть решен однозначно. Существенно, что анаэроб ные инфузории с гидрогеносомами час то имеют близких родственников с ми тохондриями, и сейчас ясно, что эти ана эробные протисты филогенетически связаны с их аэробными предшествен никами. Таким образом, вполне вероят но, что анаэробные свободноживущие
инфузории с гидрогеносомами эволюци онировали от аэробных инфузорий с ми тохондриями. Однако есть основания полагать, что гидрогеносомы инфузорий из рубца жвачных эволюционировали независимо от гидрогеносом некоторых парабазалий, т.е. жгутиконосцев, живу щих в термитах. В каждом случае гид рогеносомы оказываются вторичным приобретением, и это служит важным критерием для оценки монофилии той или иной группы. Кроме того, присут ствие прокариотных симбионтов в ком бинации с отсутствием митохондрий и гидрогеносом является полезным при знаком для разграничения, например, парабазалий и архамеб.
Теория стадийного эндосимбиоза объясняет также и происхождение пла стид. Безмитохондриальные протисты с хлоропластами не известны, поэтому
Рис. 30. Трехмерная схема (основанная на молекулярных и морфологических данных) эво люционных преобразований фототрофных эукариот. Индивидуальные геномы клетки хо зяина и э н д о с и м б и о н т а показаны линиями: пути развития автотрофов (АР) — точками, ге теротрофных прокариот (HP) — пунктиром, гетеротрофных эукариот (НЕ) — сплошной л и нией. Уровень НЕ — это уровень гетеротрофных эукариот с двумя геномами (2), возник ш и м и в результате э н д о с и м б и о з а между примитивным эукариотом и гетеротрофным про кариотом — предшественником митохондрий . РЕ — уровень фототрофного эукариота с тремя геномами, который сформировался в результате э н д о с и м б и о з а с фототрофным прокариотом (3). НЕ + РЕ — уровень вторичного э н д о с и м б и о з а между гетеротрофным эукариотом уровня НЕ и о д н и м из фототрофных партнеров уровня РЕ, представляющих появление фототрофных организмов, таких как динофлагеллаты (D), хромисты (Chr), эв - глениды без симбионтов (Ей) и с с и м б и о н т а м и (Eus) и другие таксоны; первоначальное количество геномов 4 (+ 1 редуцированный митохондриальный геном) . НЕ + НЕ + РЕ — уровень наиболее сложных а с с о ц и а ц и й : одноклеточные организмы уровня НЕ + РЕ с и м бионты гетеротрофных эукариот, что обнаружено у цилиат (Ci и Cis), фораминифер (Fo и Fos) и радиолярий («Р.» и «R»s), без симбионтов и с н и м и , соответственно; или многокле точные животные (Со — кораллы). В таких случаях количество геномов может достигать с е м и , но обычно присутствует только 5. В пределах трех низших уровней организмы с од ноклеточной организацией независимо эволюционировали в организмы с многоклеточ ной организацией, что показано стрелками влево и вправо: многоклеточные организмы возникают в филетических линиях Biliphyta (Bi), Chlorobionta (Ch), Phaeophyta (Ph), выс ших грибов (А/В — Ascomycota/Basidiomycota) и высших Metazoa (hMz) (по Хюльсману и Хаусману).


50 Эволюция и таксономия
Первичный
эндосимбиоз |
Цианобактерия |
|
|
|
клетка-хозяин I |
Rhodophyta Glaucocystophyta Viridiplantae
Вторичный
эндосимбиоз
клетка-хозяин II
Apicom- |
Prymnesio- |
Bacillario- |
Dino- |
Crypto- |
ReticuloEugienida |
Dino- |
plexa |
monada |
phyceae |
flagellata |
monada |
filosa |
flagellata |
Третичный
эндосимбиоз
клетка-хозяин |
клетка-хозяин III |
Dinoflagellata
(с криптомонадой)
В
Рис. 3 1 . Схема, объясняющая э в о л ю ц и ю и многообразие фототрофных протистов, воз никших в результате первичного, вторичного и третичного э н д о с и м б и о з о в . Первичный
эн д о с и м б и о з : поглощение путем фагоцитоза свободноживущей цианобактерии с после
ду ю щ е й ее адаптацией, ведущей (после латерального переноса генов от симбионта в ядро

общепризнано, что только эукариотная клетка уже имеющая митохондрию мог ла приобрести хлоропласт. Пластиды генетически близки современным цианобактериям. По всей вероятности, единственная разновидность цианобактерий эволюционировала каждый раз независимым путем в настоящие двумембранные пластиды Chlorophyta, Rhodophyta и Glaucocystophyta (рис. 30). Эти хлоропласты отличаются по соста ву основных и дополнительных пигмен тов: зеленые водоросли и их потомки имеют пластиды с хлорофиллами аиЬ, красные водоросли и их потомки — с хлорофиллами а и с1. С другой сторо ны, обе группы эукариот имели одно клеточных представителей, которые оказались способны стать пластидоподобными симбионтами многих эукариотных хозяев. В частности, зеленые водоросли в лице своих потомков ста ли фотосимбионтами хлорарахниевых и эвгленовых, а красные водоросли — споровиков, криптомонад, динофлагеллат, хризомонад, примнезиомонад и других. В некоторых случаях эти связи дестабилизировались, как, например, в случае бесцветных (гетеротрофных) эв гленовых или лабиринтул, и лишь со хранившийся глазок указывает на то, что в них когда-то присутствовал эндосимбионт. Пока еще неполные данные
Эволюция одноклеточных эукариот |
51 |
о симбиотических ассоциациях между различными хозяевами и эндобионтами — от первичного эндосимбиоза ко вторичному (и даже к третичному) — суммированы на рисунке 31.
Возникновение эукариотного жгу тика стало важнейшим эволюционным приобретением. Независимо от приня тия той или иной гипотезы о происхож дении этого двигательного аппарата (путем анагенеза от простых ассоциа ций микротрубочек, используемых при кариокинезе, или, напротив, в результа те маловероятного симбиоза со спиро хетами), само по себе наличие жгутиков и различные типы их расположения на клетке служат важными признаками при поисках монофилетических таксо нов. Таким образом, если бы отсутствие жгутика сопровождалось другими при митивными особенностями (отсутстви ем диктиосом или митохондрий), то та кую конструкцию следовало бы считать исходной, но до сих пор такое сочета ние признаков в современных таксонах не найдено. Вторичное отсутствие жгу тика также служит апоморфным при знаком, особенно когда имеются другие продвинутые признаки (например, на личие пластид). В качестве примера: утрата жгутика в процессе эволюции — это признак многих групп, обладающих амебоидной подвижностью или клеточ-
' У красных водорослей обнаружен только хлорофилл а. — Прим. пер.
хозяина) к пластиде со стенкой из двух мембран, которая характерна для Viridiplantae (= Chlorobionta), Rhodophyta и Glaucocystophyta. Вторичный э н д о с и м б и о з зеленых и крас ных водорослей с гетеротрофными протистами приводит к ф о р м и р о в а н и ю фототрофных клеток с пластидами, окруженными тремя или четырьмя мембранами . Третичный с и м б и оз становится возможным после редукции собственных пластид и нового с и м б и о з а с фототрофным эукариотом (по Ш т о б е и Майру) .

52 Эволюция и таксономия
ной стенкой, и может рассматриваться как адаптация к бентосному или, по зднее в истории Земли, к наземному образу жизни.
К сходным рассуждениям прибега ют при оценке наличия или отсутствия диктиосом. Удивительно, но эта ассоци ация мембранных цистерн обнаружива ется только в клетках с митохондриями или гидрогеносомами. Резонно предпо ложить, что функциональные измене ния в эволюции аппарата Гольджи, ко торый продуцирует лизосомы, были обусловлены изменением типа питания среди одноклеточных эукариот: от осмотрофии и бактериотрофии к хищни честву Кроме того, у многих протистов элементы стенки цисты и раковинки, а также экструсомы синтезируются при помощи аппарата Гольджи. Поэтому об ладающие такой многофункциональной органеллой особи оказались способны ми вырабатывать лучшие стратегии вы живания. С другой стороны, простота строения диктиосом у многих эукариот и удивительная сложность диктиосом у
Parabasalia приводят к заключению, что эта органелла развивалась в разных так сонах независимо из каких-то прими тивных структур1.
Сравнение и оценка примитивных и продвинутых морфологических призна ков в совокупности с доступными био химическими и молекулярно-биологи- ческими данными позволяет сконст руировать филогенетическое древо, которое пока еще в определенных аспек тах гипотетично (см. рис. 33). В началь ной точке видообразования находится одноклеточный организм, который ха рактеризуется следующими основными функционально-морфологическими приобретениями: два жгутика, система внутриклеточных мембран (ядерная оболочка, шероховатый и гладкий эндоплазматический ретикулум, лизосо мы), хромосомы, митотический и мейотический аппарат из микротрубочек, си стема микрофиламентов (актомиозин) для цитокинеза, а также для выполне ния цитоскелетных функций и обеспе чения подвижности, и 70S рибосомы.
1 Трудно представить какие-то более примитивные структуры, чем аппарат Гольджи, состоящий из одиночных цистерн, как, например, у грибов. Вероятно, авторы имеют в виду образование диктиосом из отдельных цистерн, как основное направление в эволюции аппарата Гольджи. —
Прим. пер.

53
Развитие систематики
Первоочередные цели зоологиче ской систематики — установить род ственные отношения между таксонами, смоделировать исторические фазы эво люции организмов (филогения) и дать рациональное объяснение фундамен тальным процессам и причинам фило генетического развития (эволюционная биология).
Со времен Левенгука и вплоть до XXI века работы по систематике протистов осуществлялись преимущественно без выявления родственных связей; единственным доступным критерием принадлежности к одному таксону было морфологическое сходство орга низмов. Например, малый размер бак терий, одноклеточных эукариот, а так же некоторых растений и животных послужили основанием для объедине ния их в группу Animalcula, или Infu soria. Изучение строения протистов и анализ их филогении были начаты Эренбергом и Дюжарденом. Первые генеалогические деревья были постро ены под влиянием теории происхожде ния видов Чарльза Дарвина (с 1842 года), которую Геккель активно приме нил к протистам. В то время были со зданы таксономические термины, мно гие из которых используются и по на стоящее время: Polycystinea Ehrenberg, 1838; Rhizopoda von Siebold, 1845; Ciliata Perty, 1852; Flagellata Cohn, 1853; Radiolaria J. Mtiller, 1858; Suctoria Claparede et Lachmann, 1858/59; Mycetozoa de Вагу, 1859; Heliozoa Haeckel, 1866; Mastigophora Diesing, 1866; Sarcodina Schmarda, 1871; Filosea Leidy, 1879;
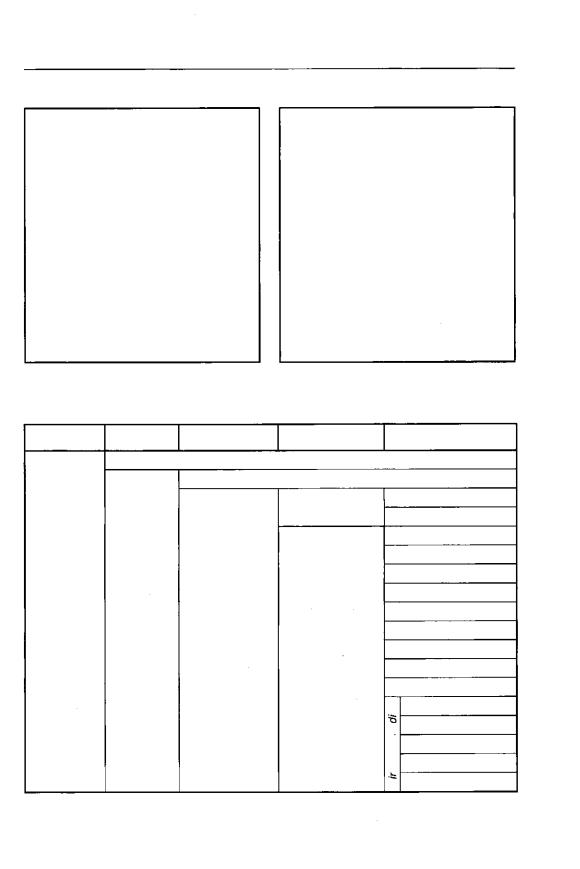
Sporozoa Leuckart, 1879; Acantharea Haeckel, 1881; Myxosporidia Biitschli, 1881. Бючли был первым, кто предло жил систему Protozoa (табл. 2).
Т а б л и ц а 2 . |
С и с т е м а Б ю ч л и |
1889 г о д а |
||
Тип |
Класс |
Подкласс |
||
|
|
|
r |
Rhizopoda |
r S a r c o d i n a - |
Heliozoa |
|||
|
|
|
|
Radiolaria |
|
|
|
|
Gregarinida |
- S p o r o z o a - |
Myxosporidia |
|||
Protozoa- |
|
|
L S a r c o s p o r i d i a |
|
- M a s t i g o p h o r a |
|
|||
- |
Infusoria |
|
|
Ciliata |
|
|
|
||
Suctoria
Благодаря ее простоте, эта схема все еще используется — преимущественно в учебниках по общей зоологии. Одна ко накопление новых знаний, включая переоценку значения признаков, под вигло Хонигберга и десять его соавто ров к созданию более современной си стемы в 1964 году. Эта схема приведе на в таблице 3.
Таблица 3. Система Хонигберга с соавторами 1964 года
Тип |
Подтип |
Класс |
||
|
|
|
|
— Sarcodina |
|
i— Sarcomasti — — Mastigophora |
|||
|
g o p h o r a |
— Opalinata |
||
|
|
|
|
|
|
— Sporozoa |
|
|
— Telosporea |
Protozoa — |
|
|
— Toxoplasmea |
|
|
|
|||
|
|
|
||
|
|
|
|
|
|
_ . |
|
|
г- Myxosporidia |
|
— C n i d o s p o r a H ... |
|||
|
|
|
|
L- Microspondia |
|
— Ciliophora |
|
— Ciliatea |
|
|
|
|||
54 Эволюция и таксономия
Ее отличия от системы Бючли за ключались преимущественно в разделе нии старого таксона Sporozoa на Cnidospora (споры с филаментом) и так назы ваемых новых Sporozoa (споры без филамента), а также в слиянии Mastigo phora и Sarcodina с образованием ново го подтипа Sarcomastigophora.
Новые ультраструктурные исследо вания в последующие десятилетия по зволили достичь нового уровня понима ния проблемы. Осознание огромного многообразия одноклеточных организ мов привело к идее о независимых фи логенетических путях развития. В 1980 году комитет по номенклатуре (Ливайн и 15 соавторов) предложил систему, в которой эта гетерогенность отражена в признании семи типов вместо типа Protozoa (табл. 4).
Т а б л и ц а 4 . С и с т е м а Л и в а й н а с с о а в т о р а м и 1 9 8 0 г о д а
П о д ц а р с т в о : P r o t o z o a
1. Тип Sarcomastigophora Honigberg & Balamuth, 1963
2. Тип Labyrinthomorpha Page, 1980
3. Тип Apicomplexa Levine, 1970
4. Тип Microspora Sprague, 1977
5. Тип Ascetospora Sprague, 1978
6. Тип Myxozoa Grasse, 1970
7. Тип Ciliophora Doflein, 1901
Опубликованный Обществом протозоологов в 1985 году «Иллюстрирован ный атлас простейших» (The Illustrated Guide to the Protozoa) содержит в подцарстве Protozoa только 6 типов с 28 классами (табл. 5). Новые по сравнению с системой Ливайна с соавторами (1980) таксоны приведены здесь в круглых скобках, а классы с новыми названиями отмечены квадратными скобками.
«Руководство по протестам» (Hand book of Protoctista), опубликованное в 1990 году Маргулис, Корлисом, Мелконианом и Чапменом, насчитывает в пре делах Protoctista, наряду с двумя сомни тельными, 35 более или менее реаль ных типов (табл. 6). В список типов вошли новообразованные таксоны, раз битые на части прежние типы Protozoa, классические полцарства и отделы во дорослей (Rhodophyta, Phaeophyta, Chlorophyta) и некоторых грибов (Hyphochytridiomycota, Plasmodiophoromycota, Chytridiomycota и Oomycota). Эта (не формальная) система уже отчасти фило генетически ориентирована, но основа на просто на наличии жгутика или при сутствии/отсутствии сложных жизнен ных циклов.
Авторы системы подчеркивают ге терогенность одноклеточных существ введением непривычно большого коли чества таксонов высокого ранга. Была проведена также радикальная ревизия хорошо известных групп. В особенной степени это коснулось прежних Rhi zopoda, которые были разделены на де вять типов. С другой стороны, неко торые таксоны мало изменились, на пример Ciliophora и преемники Spo rozoa.
Во втором издании нашего пособия (1996), используя, помимо морфологи ческих, молекулярные данные и эволю ционные гипотезы того времени, мы предложили систему простейших, под черкивающую филогенетические связи (табл. 7). Система Кавалье-Смита (1998) и схема, представленная во втором из дании «Иллюстрированного атласа про стейших» (The Illustrated Guide to the Protozoa, 2000), каждая из которых от-

|
|
Развитие |
систематики |
55 |
|
Т а б л и ц а 5 . С и с т е м а , п р и н я т а я в п е р в о м и з д а н и и « И л л ю с т р и р о в а н н о г о а т л а с а |
|
||||
п р о с т е й ш и х » 1 9 8 5 г о д а |
|
|
|
|
|
П о д ц а р с т в о P r o t o z o a (Goldfufl, |
1 8 1 8 ) v o n S i e b o l d , |
1 8 4 6 |
|
||
Тип Sarcomastigophora Honigberg & Balamuth, 1963 |
|
|
|
||
Подтип Mastigophora Diesing, |
1866 |
|
|
|
|
Класс Phytomastigophorea Calkins, 1909 |
|
|
|
||
Класс Zoomastigophorea Calkins, 1909 |
|
|
|
||
Подтип Opalinata Corliss & Balamuth, 1963 |
|
|
|
||
Класс Opalinatea Wenyon, |
1926 |
|
|
|
|
Подтип Sarcodina Schmarda, |
1871 |
|
|
|
|
Надкласс Rhizopodea von Siebold, 1845 |
|
|
|
||
Класс Lobosea Carpenter, |
1861 |
|
|
|
|
Класс Acarpomyxea Page, |
1976 |
|
|
|
|
(Класс Acrasea Schroter, 1886) |
|
|
|
||
Класс Mycetozoea de Bary, 1859 |
|
|
|
||
(Класс Plasmodiophorea Cook, 1928) |
|
|
|
||
Класс Filosea Leidy, 1879 |
|
|
|
|
|
Класс Granuloreticulosea de Saedeler, 1934 |
|
|
|
||
(Класс Xenophyophorea Schulze, 1904) |
|
|
|
||
Надкласс Actinopodea Calkins, |
1902 |
|
|
|
|
Класс Acantharea Haeckel, |
1881 |
|
|
|
|
Класс Polycystina (Ehrenberg, 1838) Riedel, 1967 |
|
||||
Класс Phaeodarea Haeckel, |
1879 |
|
|
|
|
Класс Heliozoea Haeckel, 1866 |
|
|
|
||
Тип Labyrinthomorpha Page, 1980 |
|
|
|
|
|
(Класс Labyrinthulea Levine & Corliss, 1963) |
|
|
|
||
Тип Apicomplexa Levine, 1970 |
|
|
|
|
|
Класс Perkinsida Levine, 1978 |
|
|
|
||
Класс Sporozoasida Leuckart, 1879 |
|
|
|
||
Тип Microspora Sprague, 1969 |
|
|
|
|
|
Класс Metchnikovellidea Weiser, 1977 |
|
|
|
||
[Класс Rudimicrosporea Sprague, 1977] |
|
|
|
||
Класс Microsporididea Corliss & Levine, 1963 |
|
|
|
||
[Класс Microsporea Delphy, 1963] |
|
|
|
||
Тип Myxozoa Grasse, 1970 |
|
|
|
|
|
Класс Myxosporea Butschli, |
1881 |
|
|
|
|
Класс Actinosporea Noble, |
1980 |
|
|
|
|
Тип Ciliophora Doflein, 1901 |
|
|
|
|
|
Подтип Postciliodesmatophora Gerassimova & Seravin, |
1976 |
|
|||
Класс Karyorelictea Corliss, |
1974 |
|
|
|
|
Класс Spirotrichea Butschli, |
1889 |
|
|
|
|
Подтип Rhabdophora Small, 1976 |
|
|
|
|
|
Класс Prostomatea Schewiakoff, 1896 |
|
|
|
||
Класс Litostomatea Small & Lynn, 1981 |
|
|
|
||
Подтип Cyrtophora Small, 1976 |
|
|
|
|
|
Класс Phyllopharyngea de Puytorac et al., 1974 |
|
|
|||
Класс Nassophorea Small & Lynn, 1981 |
|
|
|
||
Класс Oligohymenophorea de Puytorac et al., |
1974 |
|
|||
Класс Colpodea de Puytorac et al., 1974 |
|
|
|
||

56 Эволюция и таксономия
Т а б л и ц а 6 . С х е м а М а р г у л и с с с о а в т о р а м и
Ц а р с т в о P r o t o c t i s t a ( H o g g , 1 8 6 1 ) M a r g u l i s & S c h w a r t z , 1 9 8 8
Тип |
Класс |
С е к ц и я |
I |
Типы без жгутиковых стадий и сложных жизненных циклов
1. Rhizopoda
Lobosea
Filosea
2. Haplosporidia
Haplosporea
3. Paramyxea
Paramyxidea
Marteiliidea
4. Myxozoa
Myxosporea
Actinosporea
5. Microspora
Rudimicrosporea
Microsporea
С е к ц и я II
Типы без жгутиковых стадий, но со сложными жизненными циклами
6. Acrasea
Acrasida
7. Dictyostelida
Dictyostelida
8. Rhodophyta
Rhodophyceae
9. Conjugatophyta
Conjugatophyceae
С е к ц и я III
Типы со жгутиковыми стадиями, но без сложных жизненных циклов
10.Xenophyophora Psamminida Stanomida
11 . Cryptophyta Cryptophyceae
12.Glaucocystophyta
Glaucocystophycea
13. Karyoblastea
Karyoblastea
14. Zoomastigina
A m o e b o m a s t i g o t a
1990 г о д а
Bicoecida
Choanomastigota
Diplomonadida
Pseudociliata
Kinetoplastida
Opalinata
Proteromonadida
Parabasalia
Retortamonadida
Pyrsonymphida
15. Euglenida
Euglenophyceae
16. Chlorarachnida
Chlorarachniophyceae
17.Prymnesiophyta Prymnesiophyceae
18.Raphidophyta Raphidophyceae
19.Eustigmatophyta Eustigmatophyceae
20. Actinopoda
Polycystina
Phaeodaria
Heliozoa
Acantharia
2 1 . Hyphochytriomycota
Hyphochytrida
22. Labyrinthulomycota
Labyrinthulida
Thraustochytrida
23. Plasmodiophoromycota
Plasmodiophorida
С е к ц и я IV
Типы со жгутиковыми стадиями и сложными жизненными циклами
24 . Dinoflagellata Dinophyceae Syndiniophyceae
25. Chrysophyta Chrysophyceae Pedinellophyceae Dictyochophyceae (= Silicoflagellata)
26. Chytridiomycota Chytridiomycetes
27. Плазмодиальные слизевики Protostelida Myxomycotina
|
|
|
|
Развитие |
систематики |
57 |
Т а б л и ц а 6 . О к о н ч а н и е |
|
|
|
|
|
|
28. Ciliophora |
|
|
|
Fragilariophyceae |
|
|
Karyorelictea |
|
|
Bacillariophyceae |
|
||
Spirotrichea |
|
32 . Chlorophyta |
|
|
||
Prostomatea |
|
|
Prasinophyceae |
|
||
Litostomatea |
|
|
Chlorophyceae |
|
||
Phyllopharyngea |
|
|
Ulvophyceae |
|
|
|
Nassophorea |
|
|
Charophyceae |
|
|
|
Oligohymenophorea |
|
|
и 4 другие группы ранга порядка |
|||
Colpodea |
|
33. Oomycota |
|
|
||
29. Granuloreticulosa |
|
|
Saprolegniomycetidae |
|
||
Athalamea |
|
|
Peronosporomycetidae |
|
||
Foraminifera |
|
34. |
Xanthophyta |
|
|
|
30. Apicomplexa |
|
|
Xanthophyceae |
|
||
Gregarinia |
|
35. |
Phaeophyta |
|
|
|
Coccidia |
|
|
Phaeophyceae |
|
|
|
Haematozoa |
|
36. incertae sedis: |
|
|
||
3 1 . Bacillariophyta |
|
|
Ellobiopsida |
|
|
|
Coscinodiscophyceae |
|
Ebriida |
|
|
||
Т а б л и ц а 7. С и с т е м а Х а у с м а н а и Х ю л ь с м а н а , о п у б л и к о в а н н а я в 1 9 9 6 г о д у |
|
|||||
в о в т о р о м и з д а н и и у ч е б н и к а |
|
|
|
|
|
|
Империя |
Царство |
Подцарство |
|
Надтип |
Тип |
|
|
Microspore |
|
|
|
|
|
|
Archamoebaea |
|
|
|
|
|
|
|
|
|
Retortamonada |
|
|
|
|
|
Tetramastigota |
Axostylata |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Euglenozoa |
|
|
|
|
|
|
Heterolobosa |
|
|
|
|
|
|
Dictyostela |
|
|
|
|
|
|
Protostela |
|
Eukaryota |
|
|
|
|
Myxogastra |
|
Mastigota |
|
|
|
Chromista |
|
|
|
Dimastigota |
|
|
|
||
|
|
|
|
Alveolata |
|
|
|
|
|
Metakaryota |
|
||
|
|
|
|
|
||
|
|
|
|
Choanoflagellata |
|
|
|
|
|
|
|
Chlorophyta |
|
|
|
|
|
и |
Amoebozoa |
|
|
|
|
|
|
|
|
|
|
|
|
g> |
Granuloreticulosa |
|
|
|
|
|
w |
|
|
icertae
Actinopoda
Ascetospora
Myxozoa