

Лекция_6_ Болота_и_искуств_водоемы
.docЛекция 5.
Болота. Представляют собой неглубокие скопления воды, частично или полностью закрытые сверху растительностью. Болото – нечто переходное между водой и сушей, причем границу между заболоченными водами и заболоченной сушей провести невозможно. Характерный и обязательный признак болот – образование торфа из отмирающего мха и других гидрофильных растений. По характеру водного питания, условиям залегания и составу растительности болота подразделяют на низинные, или эвтрофные, верховые, или олиготрофные, и переходные, или мезотрофные. Низинные болота располагаются в понижениях рельефа, поверхность их вогнутая или плоская, в питании основную роль играют грунтовые воды, речные разливы, поверхностный сток и атмосферные осадки. Верховые болота находятся на повышенных формах рельефа, имеют выпуклую поверхность, питаются атмосферными осадками. Переходные болота по рассматриваемым признакам занимают промежуточное положение
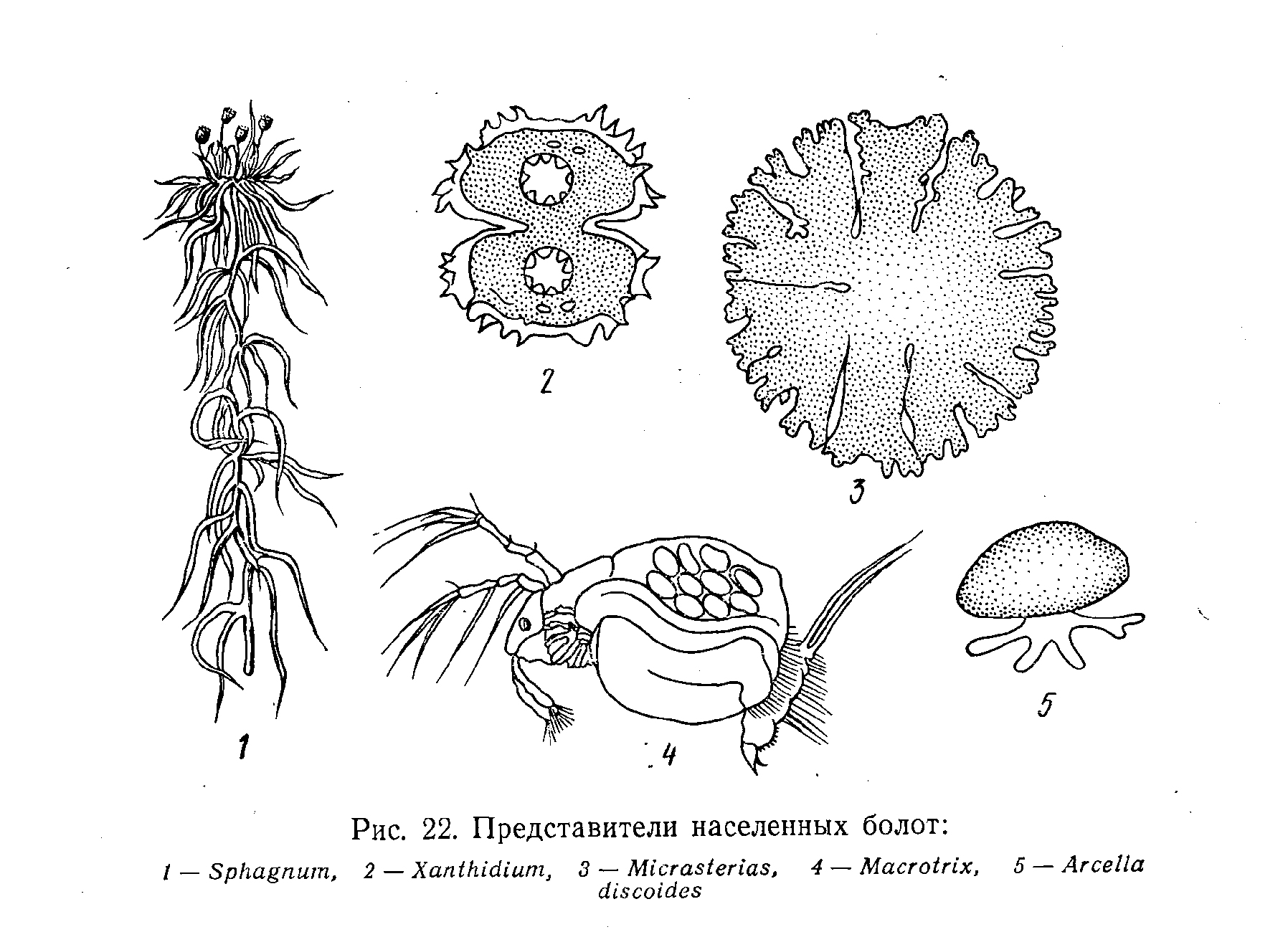
Население болот (рис. 22) отличается бедностью как по видовому составу, так и в количественном отношении. Отрицательное значение имеют обилие в воде гуминовых веществ, низкая концентрация кислорода, повышенная кислотность, особенно в верховых болотах. Во многих случаях болота бедны биогенами, в первую очередь верховые, поскольку питающие их атмосферные осадки почти не содержат солей. Поступление биогенов в толщу воды из грунта блокируется находящимися на дне отложениями торфа.
В низинных болотах нашей страны из макрофитов наиболее обычны гипновые зеленые мхи, осоки, хвощи, вейники, тростники. Для верховых болот характерны сфагновые мхи, пушица, клюква, голубика. Среди водорослей весной наиболее часто встречаются жгутиковые, , которые летом почти полностью выпадают из фитопланктона и взамен их появляются зеленые, главным образом десмидиевые; весной и осенью в воде болот в небольших количествах присутствуют диатомовые. Наиболее характерные представители зоопланктона верховых болот – коловратки, ветвистоусые (дафния, Цидорус), веслоногие и гарпактициды.
Зообентос в болотных водоемах развит слабо, из-за неблагоприятного газового режима в придонном слое воды (низкая концентрация кислорода, выделение метана, сероводорода и других ядовитых газов). Вследствие высокой кислотности воды в болотах почти не встречаются моллюски и другие животные с известковым скелетом.
Искусственные водоемы. Среди водоемов, сооружаемых человеком, наибольшее значение имеют водохранилища, пруды, судоходные и оросительные каналы. Многочисленны различного рода отстойники, дренажные сооружения, наполняемые водой рисовые чеки, всевозможные водоемы ирригационной системы.
Водохранилища. К ним относят крупные водоемы с замедленным водообменом, уровенный режим которых искусственно изменен и постоянно регулируется в целях накопления и последующего использования запасов воды. По происхождению наиболее обычны водохранилища речные, озерные и наливные. Последние сооружаются в понижениях рельефа, куда по каналам подводят избыточные паводковые воды. По конфигурации различают водохранилища вытянутые — долинные и более широкие — озеровидные; по высоте расположения — равнинные, предгорные и горные.
В зависимости от характера накопления и расхода воды водохранилища могут быть годового, сезонного, недельного и суточного регулирования. Годовые могут вмещать 20—50% годового стока реки. Водохранилища сезонного регулирования перераспределяют сток на протяжении года, накапливая воду во время паводков и срабатывая уровень в летне-зимний период. Недельное и суточное регулирование стока обычно связано с изменением норм выработки электроэнергии в рабочие и выходные дни, в светлое и темное время суток. Водохранилища длительного регулирования могут осуществлять и более короткие формы его.
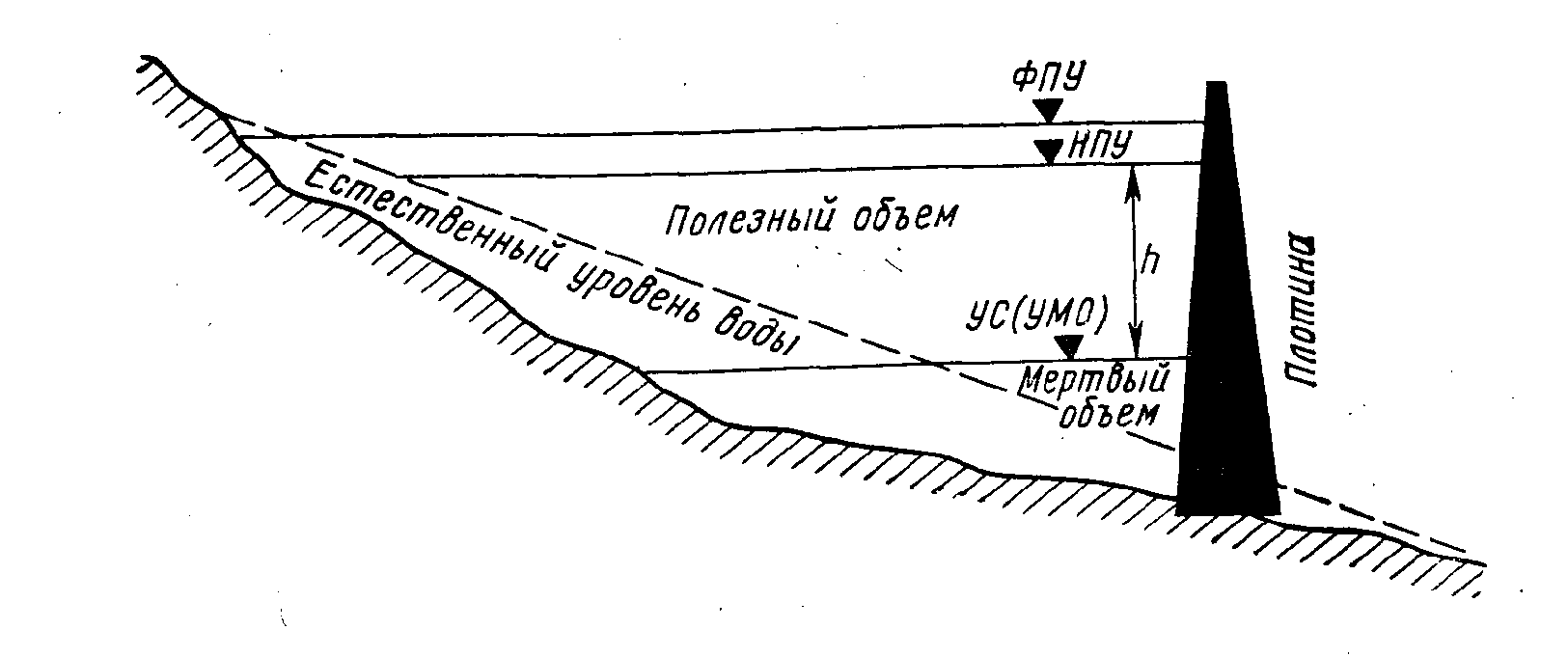
Рис. 2. Уровни и водные объемы водохранилища: ФПУ — форсированный подпорный уровень, УМО — уровень мертвого объема, НПУ—нормальный подпорный уровень (по Авакяну и Шарапову, 1977)
Весь объем водохранилища подразделяют на полезный, который может срабатываться, и мертвый, из которого попуски невозможны (рис. 2). Важные параметры водохранилища — нормальный подпорный уровень (НПУ), уровень мертвого объема (УМО) и форсированный подпорный уровень (ФПУ), допустимый на краткое время при пропуске очень высоких паводков. Котловина речных водохранилищ крайне асимметрична, поскольку повторяет очертания бывшей долины. Наибольшие глубины наблюдаются в затопленном русле; с продвижением от верхнего участка к приплотинному они возрастают.
Характерная особенность водохранилищ — частые и значительные колебания уровня, связанные с особенностями режима запасания воды и ее расхода на орошение, выработку энергии и другие нужды. Благодаря понижениям уровня большие прибрежные участки освобождаются от воды и летом высыхают, а зимой промерзают, что сопровождается гибелью очень многих обитателей прибрежья. Так как при сравнительной мелководности площадь водохранилища очень велика, во время ветров здесь наблюдается сильное волнение и энергичное размывание берегов. В связи со значительными колебаниями уровня и существованием осушной зоны сублитораль не образуется. По своим гидрологическим характеристикам речные водохранилища совмещают признаки рек и озер. С озерами их сближает значительная ширина, особенно в приплотинном участке, и как следствие этого — усиление роли ветровой деятельности; ослабление течения сопровождается осветлением воды и расслоением водной толщи — возникновением температурной и кислородной дихотомии. Сходство с рекой проявляется в сохранении течения, особенно в верхних участках и вдоль оси затопленных русел, благодаря чему гидрологические градиенты выражены гораздо слабее, чем в озерах. В верхнем участке водохранилищ гидрологический режим ближе к речному, в нижнем — к озерному.
Характерным показателем гидрологического режима служит коэффициент водообмена — отношение годового стока из водохранилища к объему последнего. В водохранилищах, построенных на равнинных реках, коэффициент водообмена, как правило, колеблется в пределах от 1 до 10. Чем он выше, тем ближе к речному режим водохранилища. На гидрологический и биологический режим водохранилищ сильно влияет сработка их уровня за счет сброса глубинных вод, которые на протяжении вегетационного периода отличаются пониженными температурами, сравнительно низким содержанием кислорода и высокой концентрацией биогенов. Вследствие сброса этих вод в водохранилище происходит накопление тепла, обеднение биогенами и улучшение кислородных условий у дна. Одновременно это влечет за собой понижение температуры, ухудшение кислородного режима и обогащение биогенами реки ниже плотины.
По видовому составу и количественному богатству население водохранилищ занимает как бы промежуточное положение между речным и озерным. В водохранилищах речного типа в верхнем участке сохраняются речные условия и речное население, в средней части флора и фауна носят промежуточный характер, а в приплотинной зоне приобретают озерные черты. В водохранилищах озерного типа население по своему составу приближается к озерному. На первых стадиях существования водохранилищ их население близко к обитателям исходного водоема. В дальнейшем оно приобретает специфический облик, зависящий главным образом от географического положения водоема.
Процесс формирования фауны водохранилища на больших равнинных реках проходит три стадии. Первая из них — разрушение существовавших до затопления реофильных, фитофильных и других группировок организмов и заселение затопленной суши и толщи воды экологически разнородным населением. Вторая стадия – образование временных группировок: в бентосе — массовое заселение в первое же лето всего дна водохранилища личинками хирономид, в зоопланктоне — массовое появление рачков и коловраток. Третья стадия формирования бентоса приурочена к завершению распространения по затопленной суше гомотопной фауны, что в основном наступает через 3—4 года после затопления и сопровождается сильным снижением биомассы бентоса. Для третьей стадии характерно уменьшение видового разнообразия зоопланктона.
Планктон сформировавшихся водохранилищ состоит в основном из бактерий, синезеленых, диатомовых и зеленых водорослей, инфузорий, коловраток и ракообразных. Бактерий содержится несколько больше, чем в реках.
Богатство фитопланктона в водохранилищах в сильной мере зависит от степени мутности поступающей воды и быстроты ее осветления. В верховьях водохранилищ, где мутность воды значительна, фитопланктона меньше, и он по своему видовому составу (преобладание диатомовых) ближе к речному, чем в средней части. В приплотинных участках фитопланктона тоже мало, так как из-за больших глубин биогены, содержащиеся в опускающихся на дно водорослях и животных, выходят из круговорота, задерживаясь в грунте, и недостаток питательных солей ограничивает развитие водорослей. В фототрофном планктоне наших водохранилищ наиболее обычны диатомовые Melosira и Asterionella, синезеленые Anabaena, Aphatiizomenon и Microcystis, зеленые Eudorina, Pediastrum и Chlamydomonas. В холодное время года возрастает роль диатомовых, летом — синезеленых.
Зоопланктон водохранилищ преимущественно представлен бесцветными жгутиковыми, инфузориями, коловратками, ветвистоусыми и веслоногими рачками, а в бассейнах рек южных морей в нем много велигеров моллюска Dreissena polymorpha. В 1 л воды бесцветных жгутиковых обычно встречается несколько миллионов, инфузорий— десятки тысяч, коловраток и ракообразных — единицы и десятки экземпляров. В верховьях водохранилищ, где речной режим выражен сильнее, преобладают коловратки, с продвижением к плотине все большее значение приобретают ракообразные. В годы с высоким уровнем воды относительное значение коловраток повышается, ракообразных — снижается. В водохранилищах с мутной водой условия жизни зоопланктона резко ухудшаются, и иногда он может существовать только в приплотинном участке или даже вовсе отсутствует. После штормов и сильных ветров вдоль берегов водохранилищ тянутся широкие полосы мутной от взвесей воды, почти лишенной зоопланктона.
Значительная гибель зоопланктона наблюдается после прохождения турбин ГЭС и пребывания в турбулентном потоке нижнего бьефа. Например, в 12 км ниже плотины Иваньковской ГЭС остается всего 15% ветвистоусых и около 35% суммарной численности зоопланктона (Дзюбан, Ривьер, 1976).
Бентос и перифитон обильны в водохранилищах, образованных на реках с прозрачной водой, и значительно беднее, когда вода мутная и дно засыпается осадками в результате выпадения минеральной взвеси. В мелких водохранилищах, занимающих обширные площади, условия существования бентоса могут резко ухудшаться из-за неустойчивости грунта, так как нередко он сильно взмучивается во время ветрового перемешивания воды.
Количество микроорганизмов в 1 г сырого ила водохранилищ колеблется в пределах 0,5—2,5 млрд. клеток.
Фитобентос представлен преимущественно прибрежными зарослями, среди которых наиболее обычны гречиха водяная, рдесты, камыш, тростник, осоки, рогоз, кувшинка и кубышка. В мутноводных водохранилищах прибрежные заросли представлены слабо из-за недостатка света и неустойчивости грунтов. Мало прибрежных макрофитов и там, где сработки уровня воды значительны и носят резкий характер, что ведет к длительному обсыханию осушной зоны и практически полной гибели находящихся здесь гидрофитов.
Зообентос в отличие от речного характеризуется значительно большей ролью вторичноводных организмов, представленных главным образом личинками насекомых, в частности хирономид. Преобладают педофилы, менее требовательные к кислороду, и только в верховьях водохранилищ встречаются настоящие реофильные формы. Соответственно этому с продвижением из верховьев к приплотинному участку видовое разнообразие донного населения снижается вследствие выпадения реофильных форм, однако в связи с возрастающим заилением грунтов биомасса бентоса не только не уменьшается, но даже заметно повышается.
Например, в верховье Волгоградского водохранилища биомасса донных животных в 2—3 раза ниже, чем в районе Саратова, а здесь она меньше, чем в приплотинном участке. На существование зообентоса в прибрежной зоне водохранилищ неблагоприятно сказывается ее периодическое обсыхание и промерзание.
Состав донного населения водохранилищ сильно зависит от того, каким он был в исходном водоеме. Как правило, наибольшее значение в бентосе имеют инфузории и другие представители микрозообентоса, личинки комаров Chironomus, Glyptotendipes и Cryptochironomus, олигохеты Limnodrilus и Tubifex, моллюски Anodonta, Unio и Viviparus. Наиболее обычные представители зообентоса водохранилищ наших южных рек — бокоплавы, мизиды и особенно моллюск Dreissena polymorpha, поскольку при заселении твердых субстратов он не встречает значительной конкуренции со стороны других лито- и фитореофильных форм, исчезающих после зарегулирования стока рек.
Нектон практически представлен только рыбами, среди которых исключительное или почти исключительное значение имеют жилые. Из их состава после зарегулирования рек реофильные формы исчезают, лимнофильные, наоборот, становятся многочисленнее. Особенно благоприятные условия для многих лимнофильных рыб складываются в первые годы существования водохранилища, когда большое количество залитой растительности образует дополнительные нерестилища. В последующем в результате отмирания залитых наземных растений условия икрометания для фитофильных рыб ухудшаются, и их численность несколько снижается. Помимо этого, неблагоприятно отражается на численности рыб обеднение их кормовой базы, наблюдающееся через 1 — 2 года после образования водохранилищ.
Пруды. Сооружаются для рыборазведения, водоснабжения населенных пунктов, полива полей, водопоя скота и ряда других целей.
Пруды могут быть плотинными, образующимися в результате запруживания речек и оврагов; копаными, питающимися атмосферными осадками, а также грунтовыми водами, и наливными, наполняемыми водой через специальные каналы из рек и ручьев. Во всех случаях пруды — это мелкие водоемы с небольшой площадью водного зеркала, часто спускаемые на зиму.
Население прудов отличается видовым однообразием, хотя по своей численности и биомассе оно часто богаче озерного. Ведущую роль в фауне прудов играют вторичноводные организмы, преимущественно представленные эврибионтными формами, что отражает большую неустойчивость температурного, кислородного и других факторов среды, в которой они обитают. Число бактерий в прудах может достигать нескольких десятков миллионов в 1 мл, как это наблюдается, например, в случае внесения в них органических удобрений. Автотрофный планктон в основном представлен зелеными водорослями, особенно протокковыми и синезелеными, меньшее значение имеют диатомовые. Из отдельных форм наиболее характерны для наших прудов зеленые Scenedesmus, Pedias- trum, Closterium и Cosmarium, синезеленые Aphanizomenoti, Ana- baena и Microcystis. Зоопланктон (см. рис. 20) слагается главным образом из инфузорий, коловраток, ветвистоусых и веслоногих рачков.
В прудах средней полосы наблюдается сезонная смена ветвистоусых рачков, которые весной преимущественно представлены видами Moina, Scapholeberis и Simocephalus, а позже Daphnia и Bosmina. Летом фауна ветвистоусых резко обедняется, и они могут исчезать из прудов, по-видимому, в связи с понижением численности бактерий и протококковых, которыми они в основном питаются. Численность инфузорий в прудовой воде может достигать 500 тыс. экз./л (Небрат, 1975).

Если пруды не спускные, фитобентос достигает в них значительного обилия (см. рис. 21). В зарослях макрофитов наиболее обычны тростник, рогоз, осоки, рдесты, роголистник, уруть и некоторые другие, на которых появляется богатая фитофильная фауна, состоящая из личинок насекомых, моллюсков, губок и мшанок. Особенно богата фауна мягкой растительности. Зообентос (см. рис. 21) включает преимущественно инфузорий и других представителей микрозообентоса, олигохет, моллюсков, личинок комаров, стрекоз, имаго и личинок жуков. Распределяется зообентос по ложу пруда довольно равномерно как в видовом, так и в количественном отношениях. Большинство организмов инфауны находится в поверхностном слое грунта толщиной 10-20 см, причем днем животные держатся на больших глубинах, чем ночью. Зимой донные организмы проникают в грунт глубже, чем летом. Нектон прудов представлен немногочисленными рыбами, в частности карасем, сазаном, линем. Искусственно разводятся карп, толстолобики, форель и некоторые другие.
Каналы и водоемы оросительной системы. Их сооружают в связи с перераспределением стока рек, для орошения, судоходства, транспорта питьевой воды и других целей. Вода, поступающая по каналам, должна удовлетворять различным требованиям. Например, в воде оросительных каналов важна концентрация некоторых ионов, а также их соотношение; в технических — общая минерализация и жесткость воды, в водопроводных — биологическая полноценность ее. Население каналов оказывает огромное влияние на качество воды и, кроме того, может существенно менять водопропускную способность каналов и их судоходные качества (зарастание тростником, камышом, рогозом и др.).
Оросительные системы в какой-то степени напоминают речные. В речной системе русло занимает самое нижнее положение, а придаточные водоемы располагаются выше. Наоборот, головной канал оросительной системы расположен выше других элементов. В главную реку впадают притоки, а из головного канала берут начало ответвления, уносящие воду потребителям.
В головных частях оросительных каналов вода по своим характеристикам близка к исходной (речной или иной), в дальнейшем ее температура, прозрачность и различные гидрохимические показатели меняются и соответственно формируется специфическое население. На камнях береговых откосов развиваются литореофилы, на дне по мере удаления от головного участка и снижения скорости течения псаммореофилы сменяются пелореофилами и пелофилами. Там, где вследствие замедления течения на дно выпадает большое количество взвешенного материала, у берегов появляются высшие водные растения, которые в свою очередь еще больше замедляют скорость течения. В фототрофном планктоне усиливается роль синезеленых и снижается значение диатомовых водорослей. Зоопланктон обычно становится богаче исходного, особенно за счет развития фитофильного комплекса. В бентофауне наибольшее значение приобретают личинки хирономид и других двукрылых, ручейников, поденок, моллюски, черви. Некоторые личинки своими домиками и чехликами укрепляют береговые откосы, роющие формы разрушают их. Обилие и структура населения откосов в облицованных и необлицованных каналах сильно отличаются, что в свою очередь в значительной мере определяет характер трансформации качества исходных вод. Ведущим элементом гидрологического режима, обусловливающим тот или иной облик населения каналов, является скорость течения.
Из разнообразных водоемов оросительной системы наибольшее значение имеют чеки рисовых полей, в которых вода удерживается земляными валиками высотой 30—40 см. Глубина воды в чеках обычно не превышает 12—15 см и задерживается в них на период вегетации риса. Характерная черта чеков — незначительность и равенство глубины, высокая прогреваемость воды в жаркое время суток и быстрое охлаждение ночью, так что перепады температуры достигают иногда 20—25°С и более. Крайне неустойчив и кислородный режим. В первую половину лета, когда рис еще не поднялся над поверхностью воды и насыщает ее кислородом, содержание газа может в два раза превышать нормальное. Когда рис поднимается над водой и фотосинтетическая аэрация ее резко снижается вследствие затенения, содержание кислорода становится ниже нормального. По сходным причинам содержание этого газа в воде ночью меньше, чем днем.
Фототрофный планктон рисовых чеков в период открытой поверхности воды в основном представлен десмидиевыми и диатомовыми водорослями, а после разрастания риса — конъюгатами и синезелеными, устойчивыми к затенению. На стеблях риса в большом количестве развиваются нитчатые водоросли, они же покрывают и дно чеков. Зоопланктон в основном состоит из веслоногих, ветвистоусых и ракушковых рачков, коловраток, инфузорий. Его биомасса может достигать очень высоких величин (10—50 г/м3). Бентофауна значительно беднее (инфузории, олигохеты, моллюски, личинки насекомых), богаче представлен зооперифитон. Многие личинки хирономид, типулид и ручейников, а также листоногий рачок щитень вредят посевам риса, поедая его всходы, подкалывая корешки, минируя листья. Иногда чеки могут становиться местами выплода кровососущих насекомых, в частности малярийных комаров. Во многих странах рисовые чеки дополнительно используются для выращивания рыбы и промысловых ракообразных, поскольку обладают высокой биологической продуктивностью. Например, на рисовых полях Узбекистана продукция зоопланктона за одно лето достигает 1,6 кг/м3 (Лебедева, Рахманова, 1978).
Подземные воды и их население
В жидкой фазе вода представлена в грунтах гигроскопической, пленочной, капиллярной и гравитационной (свободной). Свободная вода заполняет крупные капилляры и другие пустоты в породах и таким образом на различных глубинах формируются водоносные горизонты. Каждый такой горизонт подстилается водоупорным слоем. Гравитационные воды горизонта (рис. 24), залегающего на первом водоупорном слое, называются грунтовыми, а находящегося между двумя водоупорными слоями — межпластовыми. Грунтовые воды — безнапорные, межпластовые — могут быть безнапорными или напорными; в последнем случае они называются артезианскими. Из подземных вод наибольшее значение в качестве биотопа гидробионтов имеют грунтовые воды. К ним относятся пещерные, фреатические и интерстициальные. Первые приурочены к крупным пустотам в земной коре. Фреатические воды заполняют трещины и капилляры глубоких слоев земли, а интерстициальные — капилляры между частицами поверхностных песчанистых отложений. Распространены грунтовые воды на той или иной глубине почти повсеместно.
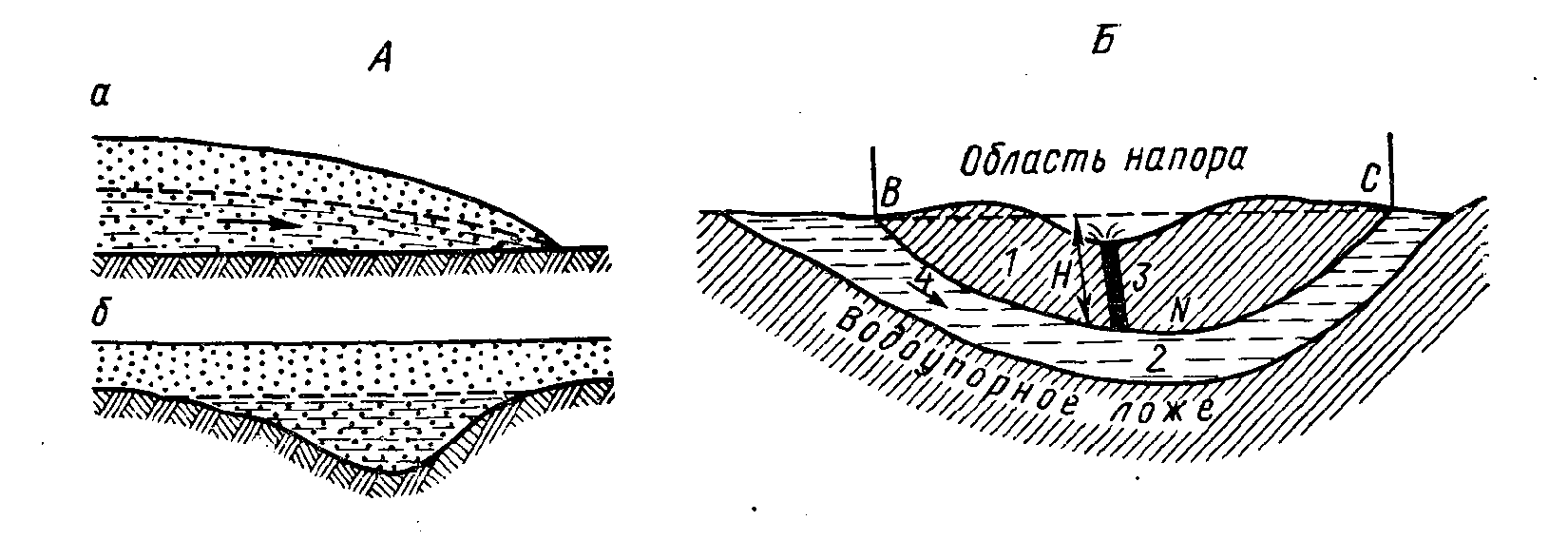
Рис. 24. Подземные воды. Схема залегания грунтовых вод (А) и схема строения артезианского бассейна
а — грунтовый поток, б — грунтовый бассейн; 1 — водонепроницаемые породы, 2 — напорный водоносный слой, 3 — скважина, 4 — направление потока; В — С — нижняя поверхность водоупорной кровли, Н — высота напора (Давыдов и др., 1973)
Обитателей подземных вод называют троглобионтами. Население артезианских вод к настоящему времени изучено очень слабо. Из грунтовых вод в гидробиологическом отношении более обследованы пещерные и интерстициальные.
Пещерные воды. В результате растворения известняков, гипса, доломитов и некоторых других горных пород в их толще возникают пустоты — пещеры, которые часто заполняет вода. Иногда она образует на дне пещер лужи или целые озера, в других случаях возникают потоки — подземные ручьи и реки. Питаются они за счет других грунтовых вод или атмосферных осадков, проникающих сквозь почву. Иногда наземная река скрывается под землю и дает начало подземному потоку, в других случаях подземный поток выходит на поверхность, образуя исток ручья или реки. Своеобразие абиотических условий пещерных вод определяет крайнюю характерность их населения.
Специфические условия, определяющие своеобразие населения пещерных вод — отсутствие света и сравнительно постоянная низкая температура. Свет в пещерах есть только вблизи входов и обычно совершенно исчезает через несколько десятков метров особенно если пустоты образуют крутые извилины. Температура пещерных вод почти постоянна на протяжении года, если они находятся в пустотах, слабо сообщающихся с атмосферным воздухом. Если пещеры хорошо вентилируются, температура их воды колеблется по сезонам, но не столь значительно, как на поверхности. Не прогреваемые солнцем пещерные воды, как правило, отличаются низкой температурой. Только в очень глубоких пещерах в связи с повышением температуры Земли (примерно ГС на 30 м глубины) водоемы могут быть теплыми.
Солевой состав подземных вод обычно отличается высокой концентрацией ионов кальция и магния, т. е. повышенной жесткостью. В водоемах Кутаисских пещер содержание кальция достигает 211 мг/л, в колодцах Саратова — 616 мг/л. Если пещерные ходы находятся в песчанистых или галечных отложениях, вода меньше минерализована. В засушливых районах Западной Сибири и Средней Азии нередко встречаются солевые пещерные воды. Газовый режим пещерных вод часто характеризуется пониженным содержанием кислорода, так как фотосинтетическая аэрация отсутствует, а проникновение газа из атмосферы нередко крайне ослаблено. С одной стороны, неподвижность воды подземных луж и озер замедляет их насыщение кислородом, с другой — не всегда его много в воздухе над пещерными водоемами.
Состав населения пещерных вод прежде всего зависит от степени их освещенности. В эвфотической зоне преобладают случайные вселенцы, троглоксены. Основной источник пищи здесь — растения и их остатки. В олигофотной зоне преобладают троглофилы, биомасса организмов здесь меньше, в значительном количестве развиваются синезеленые и бактерии. Обитатели афотической зоны — истинные троглобионты, для которых характерны стенотермность и эврифагия, представлены бактериями, простейшими, ракообразными, некоторыми моллюсками и амфибиями. Фауна состоит почти исключительно из первичноводных животных, среди которых очень много эндемичных видов. Наиболее разнообразны в видовом отношении веслоногие рачки Harpacticoida (более 120 видов), бокоплавы (более 100 видов) и креветки, сравнительно немногочисленны моллюски, единичными формами представлены коловратки, архианнелиды, пиявки и полихеты.