

Лекции по экологической физиологии / Лекция 5 ЭФ2012
.doc
Лекция 5
Эволюция газов в атмосфере
Критическое давление кислорода.
Эволюция газов в атмосфере
В присутствии кислорода питательные вещества - их углеродные кольца – могут распадаться до конечных продуктов – Н2О и СО2, и тогда энергия, заключенная в этих веществах, высвобождается. Поэтому способность окислять питательные продукты, которая появилась у живых существ в присутствии кислорода, стало эпохальным событием на Земле и привело к прогрессивной эволюция животного мира: от мелких форм к крупным, от холоднокровных к теплокровным. Такая эволюция происходила на фоне увеличения концентрации кислорода в атмосфере Земли от почти нулевых значений в начале ее существования до 21% в наше время. Ее ориентировочные величины в прошлом оценивают геологи по степени окисления земных пород разного возраста, а причиной такого повышения считают фотосинтез, осуществляемый сначала микроорганизмами, а затем и растениями, рис. 40. Одновременно концентрация СО2 в атмосфере падает, примерно, в 10 раз [44].
Принято считать, что развитие живых существ направлено от химического разнообразия и морфологической простоты к химической однородности и морфологическому разнообразию.
Примерно, первые 3 млрд. лет происходила химическая эволюция и устанавливалось биохимическое единство живых существ и лишь потом началась морфологическая эволюция, в результате которой появились разные по форме и размеру животные и растения.

Рис. 40. Развитие животного мира и эволюционное формирование сердечно-сосудистой системы [44, 75].
По оси ординат слева – шкала времени, справа - парциальное давление О2 в атмосфере.
Если сопоставить во времени изменения атмосферной концентрации кислорода и эволюцию животных организмов, то можно выделить несколько стадий [75]:
1. Примерно, 4 млрд. лет тому назад, когда концентрация О2 в атмосфере была в 1000 раз меньше современной (уровень Юри), появились первые микроорганизмы, черпающие энергию для своей жизнедеятельности из фотосинтеза. В атмосферу стал поступать биогенный кислород.
2. Через 1,5-2 млрд. лет концентрация атмосферного О2 увеличилась, примерно, в 10 раз (рО2 около 1 мм рт. ст.), и существующие тогда микроорганизмы – прокариоты начали использовать кислород как акцептор электронов, появилось дыхание. Этот уровень концентрации О2 называют именем Л. Пастера, показавшего, что с повышением рО2 в среде брожение питательных веществ меняется на их окисление. Предполагают, что случайно или нет микроорганизмы, способные к окислению веществ, поселились в более крупных анаэробных клетках и стали обеспечивать эти клетки энергией окисления, образуя для этого АТФ. Так в клетках появились митохондрии [29].
3. В дальнейшем биологическая и химическая продукции кислорода продолжалась, его концентрация в атмосфере росла. В мировом океане появились колонии клеток (теперь эукариотов), затем простейшие многоклеточные существа, у которых большая часть клеток уже не соприкасалась с внешней средой. Возникла необходимость в межорганных и внутриорганных транспортных системах газов и где-то 600 млн. лет тому назад начинается очень интересный период в эволюции животных.
Он длится всего 150-200 млн. лет, и за это время рО2 в атмосфере увеличилось немного, примерно, с 10 до 15 мм рт. ст. Однако именно в этот период формируется характерная для современных позвоночных сердечно-сосудистая система: замкнутая система трубок с О2–переносящим пигментом, О2–поглощающими капиллярами (в органах дыхания) и О2–отдающими капиллярами (в остальных органах), и с одним многокамерным сердцем, способным создавать давление, достаточное для протекания крови по 2-3 последовательно соединенным руслам.
4. Дальнейшее нарастание кислорода в атмосфере способствовало появлению озонового слоя, который стал задерживать ультрафиолетовое излучение. Поэтому живые существа выходят на сушу, сначала растения, потом насекомые и позвоночные.
Выход позвоночных на сушу (около 350 млн. лет тому назад) и возникновение легочного дыхания, а также увеличение размеров животных, еще больше отдаляют органные клетки от внешнего кислорода, хотя требования к величине рО2 на их поверхности могли оставаться прежними.
Увеличение подвижности позвоночных, а также появление теплокровности – явления, требующие повышенной скорости потребления клетками кислорода и поэтому более высокое рО2 на их поверхности, стали возможными только при нарастании концентрации О2 в атмосфере, что действительно и происходило.
Развития атмосферы Земли – повышение в ней концентрации О2 и снижение концентрации СО2.- не было линейным [86]. В истории Земли были периоды, когда эти концентрации значительно изменялись, изменялась и плотность самой атмосферы (предполагают, что часть азота замещалась более тяжелым ксеноном или легким гелием). Геофизические данные показывают, что в период 500-350 млн. лет тому назад концентрация СО2 могла быть увеличена, примерно, в 4 раза, после чего наступил период повышенной концентрации О2 (до 35 мм рт. ст. ). Он длился около 100 млн. лет и сменился периодом гипоксии (до 15 мм рт. ст.), после которого в течение примерно 100 млн. лет концентрация кислорода в атмосфере вновь увеличилась.
Предполагают, что периоды гипероксии в атмосфере совпадают с повышением ее плотности. Оба обстоятельства способствовали в период первой гипероксии появлению полета у насекомых, а во время второй гипероксии начали летать птицы. Есть антропологические свидетельства о существовании в этот период гигантских насекомых, например стрекоз с размахом крыльев более 70 см и гигантских земноводных – лягушек длиной до 2 м [86].
Критическое давление кислорода
Из всего сказанного следуют в дальнейшем иметь в виду два важных обстоятельства:
1. Дыхание (извлечение энергии из питательных веществ путем разрушения их кислородом) появилось в живых структурах (диаметром 1-2 мкм) при рО2 на их поверхности около 1 мм рт. ст. (уровень Пастера).
2. Эти структуры поселились в более крупных клетках, превратившись в митохондрии. Поэтому рО2 на поверхности этих крупных клеток должно быть более высоким, чтобы сохранить необходимый уровень рО2 на поверхности внутриклеточных митохондрий [85].
Отсюда следует, что у многоклеточных организмов системы переноса кислорода из внешней среды к органным клеткам должны были обеспечить установленные в эволюции и обязательные для окислительных процессов величины рО2 на поверхности клеток, а значит и на поверхности митохондрий.
Среди живых существ есть животные с зависимым типом дыхания (анаэробы). К ним относятся некоторые простейшие, кишечно-полостные, черви, большинство червей-паразитов, некоторые моллюски, ракообразные, насекомые.
Большая часть животных – аэробы, которые получают энергию за счет окисления питательных веществ. Они, как правило, обладают независимым типом дыхания, когда его скорость не зависит от концентрации кислорода в окружающей среде, пока она остается выше критического уровня. Величина рО2 крит. определяется кислородным запросом, и чем он выше, тем выше рО2 крит..
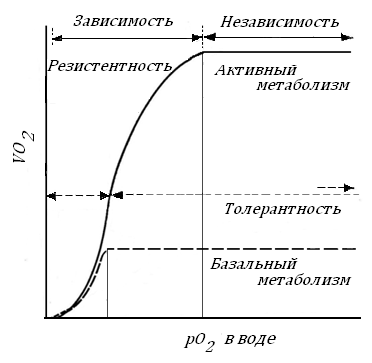
Рис. 41. Зависимость скорости потребления О2 рыбой от парциального давления его в воде в покое и при плавании (Huges, 1964), по: [41].
Показаны зоны толерантности и резистентности – неограниченной и ограниченной выживаемости. Вертикальные линии показывают критические рО2 для активного и базального метаболизма.
Рис. 41 показывает эти явления на целом животном, рыбе – гольце в условиях свободного плавания и базального метаболизма. Можно видеть зону независимого дыхания и зону – зависимого.
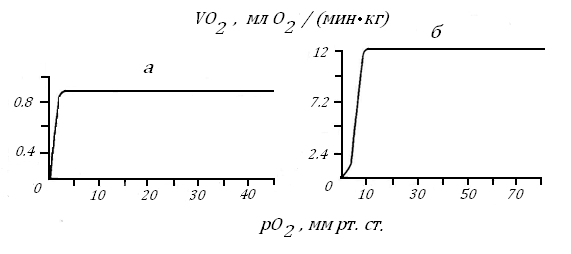
Рис. 42. Скорость потребления кислорода in vitro в зависимости от его содержания в среде волокнами белой (а) и красной (б) скелетных мышц байкальского хариуса [78].
На рис. 42 приведен второй пример с зонами независимого и зависимого типов дыхания и величинами критического рО2, но уже для изолированных волокон красной и белой скелетных мышц другой рыбы - байкальского хариуса. Скорость дыхания этих волокна различалась в 14 раз, а рО2 крит. - в 3.6 раза, табл. 17.
Для понимания условиях, в каких живет и функционирует клетка или мышечное волокно, необходимо знать величины рО2 крит для этих структур. К сожалению, этих данных мало, так как очень трудно, не повредив клетку, выделить ее из органа и уже во внеклеточной среде измерить ее рО2 крит. Больше всего таких данных получено на скелетных волокнах.
Таблица 17. Параметры дыхания изолированных мышечных волокон
зрелых позвоночных [5, 77]
|
Параметры волокна |
Курица |
Крыса |
Нерпа |
Хариус |
||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
|
Критическое рО2, мм рт. ст. |
33 |
7.7 |
38 |
26 |
17 |
16 |
8.9 |
2.5 |
|
Скорость потребления О2, млмин-1кг-1 |
18 |
4.2 |
39 |
15 |
23 |
14 |
13 |
0.9 |
|
Диаметр волокна, мкм |
67 |
67 |
65 |
76 |
53 |
54 |
75 |
150 |
Примечание. Исследовались волокна мышц: 1 и 2 - красная икроножная и белая грудная у домашней курицы, 3 и 4 – - красная четырехглавая и белая прямая бедра у лабораторной крысы, 5 и 6 – ременная с низкой концентрацией миоглобина и длиннейшая спины с его высокой концентрацией у байкальской нерпы, 7 и 8 – боковые красная поверхностная и белая большая у байкальского хариуса.
Замечу, что в табл. 17 приведены данные о рО2 крит. в мышечных волокнах, помещенных в среду с сукцинатом натрия, стимулирующим их окислительный обмен. Она показывает, что при одинаковом диаметре волокон скорость их дыхания пропорциональна величине рО2 крит (сравните 1 и 2). В более тонком волокне сравнительно высокая величина VO2 не требует столь высокого рО2 крит (сравните данных граф 3 и 4, 7 и 8). На соотношение этих двух показателей оказывает влияние концентрация в волокне миоглобина, депонирующего кислород (сравните данные граф 5 и 6). В табл. 17 можно видеть, что величины рО2 крит, для разных мышечных волокон теплокровных может колебаться от 8 до 38 мм рт. ст. Состояние кислородного голодания для этих волокон, так и для разных клеток наступает, когда концентрация кислорода в окружающей их среде становится меньше критической.
На рис. 15, который мы ранее видели, показано, что наибольший выход кислорода из оксигемоглобина у различных теплокровных происходит во время снижения рО2 в крови от 80-50 до 20-5 мм рт. ст.. Следует предполагать, что это падение рО2 как раз и должно происходить в период протекания крови по капилляру, при этом низкий уровень его должен оставаться выше, чем рО2 крит, для окружающих их паренхиматозных клеток и волокон.
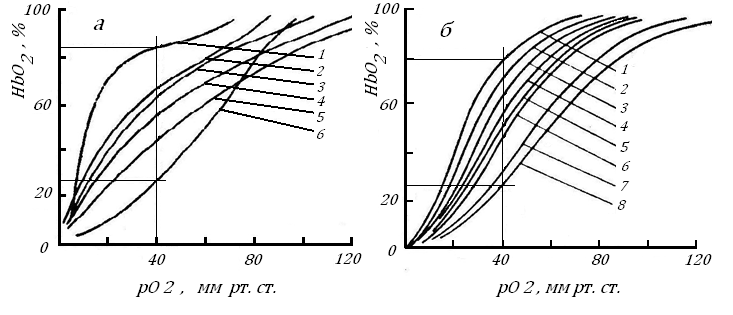
Рис. 15. Повторение Кривые диссоциации оксигемоглобина у млекопитающих с разной массой тела, а - (Филатова, 1944), по [47], б - [73].
По оси абсцисс – рО2 в среде, по оси ординат - концентрация HbO2, %. Звери на левом рисунке: 1 - лошадь, 2 - овца, 3 - степной хорек, 4 - кошка, 5 - желтый суслик, 6 - морская свинка; на правом рисунке: 1 - слон, 2 - лошадь, 3 - человек, 4 - овца, 5 - лиса, 6 - кошка, 7 - крыса, 8 – мышь.
Сложности методического характера не позволяют пока в разных органах достаточно точно измерить рО2 в капиллярах и примыкающих к ним артериолах и венулах. Эту неопределенность можно видеть на рис. 43, где приведены данные 15 исследовательских групп в период 1972-1995 гг, характеризующие рО2 в скелетных мышцах крыс, хомячков, кроликов, кошек и собак, коже хомячков, кишечной стенке крыс и кроликов, миокарде собак и в мозговой коре крысы. Для измерения рО2 использовались различные методические приемы – микроэлектроды, микроспектрофотометрию, криоскопию и фосфоресценцию
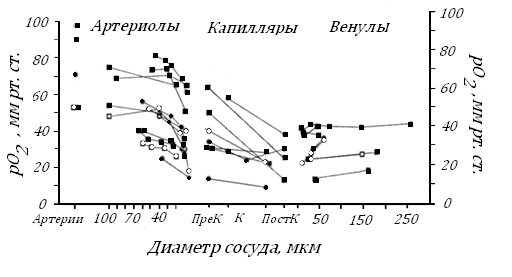
Рис. 43. Величины рО2 в плазме кровеносных сосудах различных органов у лабораторных животных, измеренных исследователями в период 1972-1995 гг [104].
Подробности о животных и органах см. в тексте. Соответствующие знаки и соединяющие линии показывают данные одной исследовательской группы. ПреК, К, ПостК – прекапилляры, капилляры и посткапилляры
Среди довольно разрозненных величин рО2 в микрососудах на рис. 43 большая часть из них скорее иллюстрирует методические сложности его измерения. Однако некоторые данные, полученные на мышцах хомячков, кошек, собак, миокарде собак и крыс и на мозговой коре кошек [103], четко показывают: наибольшее падение рО2 в капиллярном отделе (от 70-40 до 40-15 мм рт. ст.) соответствует самому резкому снижению кривой диссоциации оксигемоглобина. Похожие данные были получены в сердце крыс и мозговой коре кошек [104].
Завершая раздел о величинах рО2 крит, еще раз замечу, что скорости окислительных процессов и рО2 крит, в клетках пока исследованы недостаточно. Остается неизвестным, какие рО2 крит могут быть на поверхности клеток, живущих в регионах с низким рО2, например, клеток печени, получающих кислород, в основном, из воротной вены. Неясно, какова величина рО2 крит у макрофагов, которые до 95% потребляемого ими кислорода уже на своей поверхности превращают в радикалы и используют их для уничтожения вредных или ненужных структур и организмов.
Совсем мало данных о величинах рО2 внутри самих клеток. Например, оказалось, что в мышечном волокне его величина (она измерена по фосфоресценции порфироновых соединений в волокне), изначально близкая к рО2 во внешней среде, при сокращении волокна значительно снижается, рис. 44.
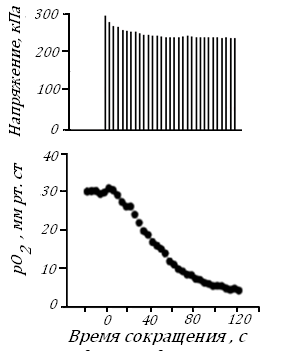
Рис. 44. Мышечное напряжение (вверху) и внутриклеточное рО2 одиночного мышечного волокна лягушки в покое и во время 2-минутного тетанического изометрического сокращения при раздражении электрическим током [94].