

Lektsii_po_biofizike_Baskakov_Kapilevich
.pdfАктивность ПК-С не зависит от кальмодулина, но многие агенты, которые известны как антагонисты КМ, ингибируют фермент. В их число входят такие вещества, как трифторперазин, хлорпромазин, кальмидозолиум и соединение W-7. Ингибирующее влияние на ПК-С оказывают флуфеназин, галоперидол, хлопротиксен, имипрамин, местные анестетики (дибукаин и тетракин), верапамил, фентоламин, адриамицин, гепарин, полимиксин В, витамин Е. Ингибирующее влияние этих агентов не обусловлено их взаимодействием с активным центром ПК-С: каталитически активный фрагмент фермента, полученный при ограниченном протеолизе, не чувствителен ни к одному из них. Кинетический анализ свидетельствует, что эти вещества конкурентно ингибируют связывание ПК-С с фосфолипидами и не влияют на взаимодействие с ионами кальция или ДАГ. Обладая высоким сродством к фосфатдилсерину и фосфатидилэтаноламину, они ингибируют ПК-С, модифицируя гидрофобные взаимодействия между ферментом и фосфолипидами.
Большой прогресс в изучении роли ПК-С в регуляции клеточных функций был достигнут благодаря использованию форболовых эфиров.
Форболовые эфиры (ФЭ), промоторы опухолей, способны замещать ДАГ и прямо активировать ПК-С в присутствии ионов кальция и фосфатидилсерина. Эти соединения, подобно ДАГ, повышают сродство фермента к кофакторам. Форболовый эфир, ТФА (12-0-тетрадеканоил-13-форболацетат) полностью активирует ПК-С при концентрации ионов кальция 0,1 мкМ. Для активации ПК-С присутствие фосфатидилсерина обязательно, но другие фосфолипиды могут проявлять с ним положительную (фосфатидилэтаноламин) или отрицательную (фосфатидилхолин, сфингомиелин) кооперативность.
В отличие от ДАГ, транзиторно образующегося в мембране и быстро деградирующего, ФЭ, встраиваясь в мембрану, действуют длительное время, так как дитерпены метаболизируются очень медленно.
Активация ПК-С во многих тканях изменяет состояние ионтранспортирующих систем, модулирует чувствительность молекулярных механизмов к другим вторичным мессенджерам, модифицирует мембранные рецепторы, изменяет активность метаболических процессов.
В настоящем разделе пособия нами рассмотрены различные пути внутриклеточной сигнализации. Следует заметить, что в одной и той же клетке присутствуют различные сигнальные системы. Таким образом, изменение активности клетки – результат сложных взаимоотношений между собой сигнальных путей, имеющихся в данной клетке.
101
Тест-задания
1.Клеточные мембраны выполняют следующие функции:
A.Компартментация
B.Рецепторная
C.Транспортная
D.Проведение нервного импульса
E.Мышечное сокращение
F.Межклеточные взаимодействия
2.В состав биомембран входят:
A.Белки
B.Липиды
C.Гликопротеиды
D.Вода
3.Мембранные липиды представлены следующими классами:
A.Фосфолипилы
B.Гликопротеиды
C.Гликолипиды
D.Стероиды
4.К группе фосфолипидов относятся:
A.Фосфатидилхолин
B.Фосфотидилинозит
C.Холестерин
D.Сфингомиелин
E.Гликофорин
F.Фосфоламбан
G.Фосфатидилсерин
5.Наибольшее влияние на микровязкость мембран оказывают:
A.Холестерин
B.Фосфолипиды
C.Цереброзиды
D.Сфингомиелин
6.Свойство амфифильности липидных молекул заключается в следующем:
A.Вокруг них формируется гидратная оболочка
B.Они способны образовывать конгломераты
C.Они содержат полярные и неполярные группировки
7.Липидные молекулы являются:
A.Гидрофобными соединениями
B.Гидрофильными соединениями
C.Амфифильными соединениями
8.Липиды в водной среде образуют следующие виды структур:
A.Ламмелярные
B.Везикулярные
C.Мицеллярные
D.Капиллярные
9.Жидкокристаллическое состояние мембранных липидов характеризуется следующими свойствами:
A.Лиотропный мезоморфизм
102
B.Термотропный мезоморфизм
C.Анизотропия
D.Изотропия
10.Фазовые переходы в мембране осуществляются между следующими состояниями:
A.Гель – золь
B.Гель – жидкая фаза
C.Жидкий кристалл – гель
11.Кооперативность фазовых переходов наблюдается:
A.В однородных мембранах
B.В неоднородных мембранах
C.В везикулах
12.Выделяют следующие механизмы генерализации фазовых переходов в мембране:
A.Энтальпийно-энтропийная волна
B.Активная волна преобразований
C.Первично-диффузионная генерализация
D.Вторично-диффузионная генерализация
13.К мембранным белкам относятся:
A.Ферментативные
B.Структурные
C.Сократительные
D.Рецепторные
14.По расположению в мембране белки делятся на:
A.Переферические
B.Интегральные
C.Ферментативные
D.Полуинтегральные
E.Монотопические
15.Белки мембран составляют следующие функциональные группы (укажите неправильный ответ):
A.Ферментативные
B.Белки цитоскелета
C.Сократительные
D.Рецепторы
16.Липиды могут модифицировать структуру мембранных белков:
A.Вторичную
B.Третичную
C.Четвертичную
17.Углеводы в мембранах присутствуют в виде:
A.Гликопротеинов
B.Протеогликанов
C.Гликолипидов
D.Полисахаридов
18.Углеводы в мембране выполняют следующие функции:
A.Транспортная
B.Рецепторная
103
C.Окислительная
D.Межклеточные контакты
19.Мозаичную модель мембраны предложили:
A.Синжер и Николсон
B.Даниэли и Девсон
C.Варбург и Нигилейн
D.Гортер и Грендель
20.Современным представлениям о структуре мембран соответствует:
A.Модель липидного бислоя
B.Мозаичная модель
C.Унитарная модель
Правильные варианты ответов: |
|
|
|
|
|
1. |
A, B, C, D, F |
8. |
A, B, C |
15. |
A, B, D |
2. |
A, B, C |
9. |
A, B, C |
16. |
B, C |
3. |
A, C, D |
10. |
C |
17. |
A, B, C |
4. |
A, B, D, G |
11. |
A |
18. |
B, D |
5. |
A |
12. |
A, B, D |
19. |
A |
6. |
C |
13. |
A, B, D |
20. |
B |
7. |
C |
14. |
A, B, D, E |
|
|
104
VII. ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНЫ
Введение
Биологические структуры относятся к открытым термодинамическим системам, постоянно обменивающимся в течение жизни с окружающей средой не только энергией, но и веществом. Роль биологических мембран – сопряжение и регуляция потоков энергии, вызывающих и сопровождающих процесс транспорта. Поэтому транспорт веществ через биологические мембраны – необходимое условие для существования жизни. Нарушение мембранного транспорта ведет к возникновению различного типа клеточных дисфункций и патологий.
Классификация видов транспорта
Различают несколько классификаций видов транспорта:
1.Относительно затрат энергии в процессе переноса веществ транспорт подразделяется на активный и пассивный.
Активный транспорт осуществляется с затратой энергии (за счёт гидролиза АТФ) и против градиента концентрации (иногда данный вид транспорта называют первично-активным).
Если же перенос веществ обеспечивает энергия, обусловленная градиентом других ионов, такой транспорт называют сопряженным (иногда используется термин вторично-активный транспорт).
Пассивный транспорт осуществляется без затраты энергии, протекает самопроизвольно и по градиенту концентрации. Градиент в процессе
пассивного транспорта снижается).
2.Относительно сопряженности переноса веществ через мембрану выделяют:
Унипорт – перенос данного вещества (иона) не зависит от наличия и переноса других веществ и котранспорт – перенос данного вещества (иона) зависит от наличия и переноса других веществ. В этом случае скорость суммарного процесса контролируется наличием и доступностью для систем переноса обоих партнеров транспортного процесса (рис. 32).
Котранспорт относительно направления подразделяют на симпорт и антипорт. При симпорте осуществляется однонаправленный и одновременный
транспорт, а при антипорте – одновременный, но разнонаправленный перенос веществ.
105

Рис. 32. Виды транспорта.
3. Специфические механизмы переноса веществ, связанные с нарушением структурной целостности мембраны: эндоцитоз и экзоцитоз – важнейшие процессы жизнедеятельности клетки, механизмы высвобождения медиатора в синаптическую щель, фагоцитоз лейкоцитов и др.
Методы изучения транспорта
1.Осмотический – изменение объёма клетки при транспорте воды.
2.Химический (цитохимический) – проникновение красителей, постановка цветных реакций.
3.Биохимический – функция ферментов – участников транспорта.
4.Изотопных меток: углерода (С14), натрия (Na22), рубидия (Rb86) и др.
5.Спектро- и флуорометрический – изменение параметров поглощения и флуоресценции специфических веществ-меток, меняющих свои характеристики после взаимодействия с внутриклеточными структурами.
6.Опосредованные методы – измерение рН, вольтамперных характеристик.
Пассивный транспорт и его виды
Подчиняется закону Фика:
dm DS dc dc dx .
Скорость переноса вещества зависит от градиента концентрации (dc/dx), площади (S) и коэффициента (D) диффузии.
Подразделяется на (рис. 33):
1.Простую диффузию.
2.Облегченную диффузию.
106

Рис. 33. Зависимость скорости пассивного транспорта от концентрационного градиента
1 – простая диффузия;
2 – облегченная диффузия.
Простая диффузия отличается отсутствием специальных систем для переноса веществ. Подчиняется правилам Овертона: скорость проникновения через мембрану прямо пропорциональна липотропности вещества и обратно пропорциональна его размерам.
Позднее Колландер из уравнения Фика вывел более удобно выражение для расчета потока (J) поступления веществ в клетку:
J P(c1 c2 ), где
с1 и с2 – разность концентраций, Р – коэффициент проницаемости мембрвны для данного вещества.
Облегченную диффузию отличает наличие специфических структур, осущетсвляющих перенос вещества через мембрану.
Наличие переносчиков изменяет кинетику транспорта, и она становится сходной с уравнениями ферментативного катализа, только в роли фермента выступает переносчик, а в роли субстрата – переносимое вещество (S):
Js Jmax S
KT S , где
КT – константа транспорта соответствует константе Михаэлиса и равна концентрации S при Js=Jmax/2.
Представители систем облегченной диффузии ионов (симпорт, антипорт) с участием специфических переносчиков:
Na+/Cl—обмен |
|
Na+/Na+--обмен |
|
Na+/H+ – обмен |
антипорт – котранспорт |
Na+/Ca2+-обмен |
|
Cl-/HCO3-+-обмен |
|
Cl-/ Cl--обмен |
|
K+/Cl—и Na+,K+,2Cl- – симпорт – котранспорт
107
Унипорт
Ионофоры – небольшие гидрофобные молекулы, которые растворяются в липидном слое мембран и увеличивают проницаемость мембран для ионов. Большинство ионофоров синтезируется бактериями (в качестве оружия против своих конкурентов), некоторые являются антибиотиками. Широко используются в клеточной биологии.
Ионофоры делятся на:
1.Подвижные переносчики. Имеют кольцеобразную структуру, и контактируют с липидным слоем: наружная часть – гидрофобная; внутренняя часть – гидрофильная содержит карбоксильные атомы кислорода.
2.Каналообразующие. Состоят из линейных полипептидов, которые
имеют гидрофобные боковые цепи. Две таких молекулы образуют канал.
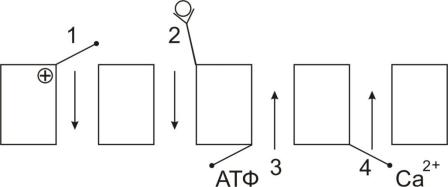
Ионные каналы (унипорт) классифицируют :
А) по типу ионов: натриевые, калиевые, кальциевые и хлорные каналы; Б) по способу регуляции (Рис. 34):
1)потенциал-чувствительные
2)хемочувствительные (рецептор-управляемые)
3)внутриклеточными веществами (ионами, ФАВ).
Рис. 34. Классификация ионных каналов по способу регуляции 1 – потенциал-чувствительные; 2 – хемочувствительные (рецептор-управляемые); 3 – АТФ-чувствительные; 4 – кальций-зависимые.
Натриевые и калиевые каналы
Размеры ионных каналов меньше одного нанометра (0,2-0,4 нм). Эти структуры обеспечивают важнейшие электрофизиологические свойства клеток и тканей, обеспечивая прохождение в 1 с 106-109 ионов, регулируются мембранным потенциалом и БАВ. Через ионные каналы Na+ и K+ следуют совместно с одной молекулой воды – гидратной оболочкой, хотя в растворе у иона таких оболочек может быть несколько (до 10). Таким образом, при прохождении через ионный канал происходит дегидратация катиона, на что требуется энергия. Роль этой энергии выполняет градиент ионов, так как ионный транспорт относится к пассивному виду переноса веществ через
108
мембрану. В процессе переноса катионов должны выполняться два основных условия (фактора):
1.Стерический – совпадение размеров катиона и гидратной оболочки с размерами канала.
2.Энергетический – взаимодействие катиона с карбоксильными (отрицательно заряженными группировками самого канала).
Кальциевые каналы
Кальциевые каналы делятся на потенциал-зависимые и рецепторуправляемые. В нормальных условиях внутри клетки [Ca2+]in = 10–7 - 10–6 M.
Через кальциевый канал проникают только двухвалентные катионы в соответствии с рядом:
pKBa2+ > pKSr2+ > pKCa2+ > pKCo2+ > pKNi2+ > pKCd2+.
Селективный фильтр кальциевого канала содержит карбоксильную группу. Все двухвалентные катионы, которые связываются с этой группой слабее, чем кальций, проникают лучше (Ba+ и Sr+). Катионы, которые связываются с этой
группой сильнее, чем кальций, являются блокторами кальциевых каналов
(Co2+,Ni2+,Cd2+).
Регуляторный центр кальциевого канала располагается у внешнего устья. В бескальциевом растворе (с добавлением хелаторов Са2+ ЭГТА, ЭДТА) происходит модификация кальциевых каналов: они сохраняют потенциалзависимость, но изменяют свою селективность и становятся селективными для натрия.
Процесс инактивации кальциевых каналов более сложный, чем натриевых каналов. У натриевых каналов происходит инактивация вследствие деполяризации мембраны, у кальциевых инактивация зависит от силы кальциевого тока. Чем выше ICa, тем быстрее инактивация, которая развивается
в результате увеличения внутриклеточной концентрации ионов кальция [Ca2+]in до 10–6 М.
Проводимость кальциевых каналов подвержена также метаболической регуляции – она зависит от содержания в цитоплазме циклического АМФ.
Кальциевые каналы подразделяют на: медленные:
порог активации – 30 мВ
двухфазная инактивация
блокирование производными 1,4-дигидропиридина
быстрые:
порог активации – 70–60 мВ
быстрая инактивация
отсутствие чувствительности к 1,4-дигидропиридинам.
109

Активный транспорт
Перенос иона в процессе активного транспорта осуществляется транспортными АТФ-фазами, энергозависимой лимитирующей стадией которого является гидролиз АТФ. Так происходит первично-активный транспорта.
Если же перенос веществ обеспечивает энергия, обусловленная градиентом ионов, созданным в ходе первично-активного транспорта, то это вторично-активный транспорт. С его помощью осуществляется перенос сахаров и аминокислот за счет энергии, обусловленной градиентом ионов, чаще натрия, созданного первично-активным транспортом.
Функциональное значение активного транспорта определяется тем, что он осуществляет неравномерное распределение ионов между клеткой и средой – для большинства тканей концентрация внутриклеточного калия больше, чем внеклеточного, а концентрация внутриклеточного натрия меньше, чем внеклеточного. Поддержание постоянного ионного состава клетки обеспечивает ионный гомеостаз, необходимый для осуществления жизненно важных градиентзатратных процессов.
Транспортные АТФ-азы прокариотических и эукаритических клеток делятся на 3 типа: P-тип, V-тип, F-тип.
АТФ-азы P-типа являются примером осуществления обязательной стадии фосфорилирования и дефосфорилирования, сопряженной со структурными переходами белков, то есть образования ковалентного фосфорилированного промежуточного продукта (фосфомедиата).
К АТФ-зам цитоплазматической мембраны этого типа относятся:
Na,+K+– АТФ-аза
Ca2+–АТФ-аза плазматическая мембрана эукариот. H+– АТФ-аза
Внутриклеточные АТФ-азы P–типа:
Ca2+ – АТФ-аза эндо-(сарко) плазматического ретикулума эукариот.
К+ – АТФ-аза наружных мембран прокариот. Устроены довольно просто, действуют как насос.
АТФ-азы V-типа находятся в мембранах в вакуолях дрожжей, в лизосомах, эндосомах, секреторных гранулах животных клеток (Н+–АТФ-азы).
АТФ-азы F-типа находятся в мембранах бактерий, в хлоропластах, митохондриях. Образованы:
1.Водорастворимой частью – F1, которая состоит из нескольких субъединиц и обладает каталитической активностью.
2.Гидрофобная часть F0, участвующей в транслокации водорода. Особенностью АТФ-азы F-типа является способность синтезировать АТФ
за счет переноса протонов по градиенту концентрации.
110