

2015-СК-біохімія-10-Обмін вуглеводів-2-ІКТ
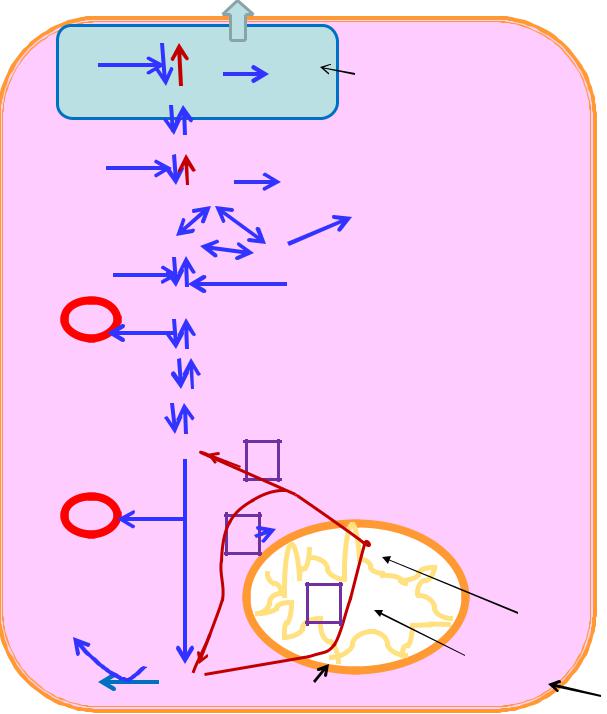
.pdfПеретворення пірувату в оксалоацетат (щавелевооцтову кислоту; щок) за дії ферменту піруваткарбоксилази (коензимна форма СО2- транспортуючого вітаміну Н, макроергічний зв'язок АТФ витрачається на карбоксилювання зв'язаного з ферментом біотину)
піруват + СО2 + АТФ → оксалоацетат + АДФ + Фн
3)
Перетворення оксалоацетату в фосфоенолпіруват за дії фосфоенолпіруваткарбоксикінази (яка локалізована в цитозолі, у людини – в цитозолі, частково – в мітохондріях):
оксалоацетат + ГТФ → фосфоенолпіруват + СО2 + ГДФ
2)перетворення фруктозо-1,6-дифосфату в фруктозо-6-фосфат за дії фруктозо-1,6-дифосфатази
фруктозо-1,6-дифосфат + Н2О → фруктозо-6-фосфат + Фн;
1)перетворення глюкозо-6-фосфату в глюкозу за дії глюкозо-6- фосфатази
глюкозо-6-фосфат + Н2О → глюкоза + Фн;
11
11
АТФ
АТФ
Фн, НАД+
АТФ
глюкоза
|
|
|
|
|
ендоплазматичний |
|
|
|
1а |
|
Фн |
||
Г-6- |
|
|
ретикулум |
|||
Ф |
|
|
|
|||
Ф-6-Ф |
1а- Глюкозо-6-фосфатаза/ 1б – |
|||||
фруктозо-1,6-диФ-фосфатаза |
||||||
|
|
1б |
||||
|
|
Фн |
|
|||
|
|
|
|
|
||
Ф-1,6диФ |
ліпіди |
|||||
|
||||||
ГАФ ДОАФ
НАДН,Н+
1,3диФГ
 АТФ
АТФ
3ФГ 2 – Піруваткарбоксилаза
2ФГ
/ ФЕП-карбоксикіназа 3 – Малік-ензим
|
ФЕП |
2 |
|
цитозоль |
|
|
|
||
|
|
щок |
|
|
АТФ |
|
СО2 |
|
|
|
|
3 |
мал |
|
|
|
|
||
НАД+ |
|
АТФСО2 2 |
щок |
малат |
|
ПВК |
|
||
НАДН |
Оксалоацетат |
|
ПВК |
||
мітохондрія |
||
|
Глюконеогенез
Г-6-Ф – глюкозо-6-фосфат; Ф-6-Ф – фруктозо-6- фосфат; Ф-1,6-диФ – фруктозо-1,6-дифосфат;
ГАФ |
– |
гліцеральдегідфосфат; ДОАФ – диоксиацетонфосфат; 1,3- диФГ – 1,3- дифосфогліцерат; 3ФГ – 3- фосфогліцеринова кислота; 2-фосфогліцеринова кислота; ФЕП – фосфоенолпіруват; ПВК – піровиноградна кислота; лактат – молочна кислота; щок – щавелевооцтова кислота (оксалооцтова кислота або «оксалоацетат»); мал – яблучна кислота («малат»)
12
клітина
Сумарна реакція глюконеогенезу:
2 піруват + 4 АТФ + 2 ГТФ + 2 НАДН + 2Н+ → глюкоза + 2 НАД+ + 4 АДФ + 2 ГДФ + 6 Фн
Синтез однієї молекули глюкози з двох молекул пірувату потребує витрат 6 макроергічних зв'язків (звідси, глюконеогенез – ендергонічний процес).
13

Утворення пірувату з лактату за дії лактатдегідрогенази в
глюконеогенних тканинах (печінка, нирки), де його спрямованість визначається відповідними відношеннями НАД+/НАДН, піруват/лактат, присутністю певного ізоферменту ЛДГ.
Джерелом лактату для глюконеогенезу в печінці є лактат, який надходить через кров з місць його утворення (переважно зі скелетних м'язів).
Лактат + НАД+ → піруват + НАДН + Н+
Безпосередні попередники глюкози при її синтезі (власне, метаболіти глюконеогенезу) – це піруват, оксалоацетат, фосфоенолпіруват.
Глюкогенні амінокислоти – це ті амінокислоти, які можуть втрачати аміногрупу (в реакціях дезамінування або трансамінування), тому вони розглядаються як субстрати глюконеогенезу.
14
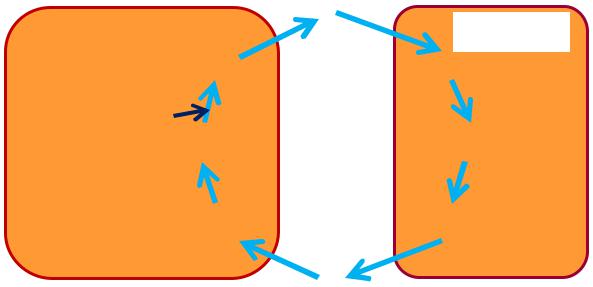
Глюкозо-лактатний цикл
Цикл Корі
У печінці |
|
||
|
|||
|
|
||
Глюконеогенез |
|
|
|
|
|
|
|
Глюкоза |
Кров |
||
6 АТФ |
|||
|
|||
Піруват |
|
||
Лактат |
|
||
|
|
|
|
Утворений активними
У м'язах Гліколіз
Глюкоза
 2 АТФ Піруват
2 АТФ Піруват
Лактат
м'язами лактат
перетворюється в глюкозу в печінці. Цей цикл зсуває частину метаболічних перетворень від активних м'язів до печінки.
15
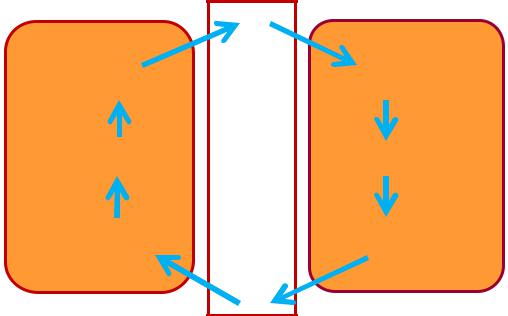
Глюкозо-аланіновий цикл
Важливий субстрат глюконеогенезу в печінці – аланін, який може утворюватися у зворотній реакції трансамінування пірувату з глутаматом у скелетних м'язах:
піруват + глутамат → L-аланін + α-кетоглутарат
Вивільняючись із працюючих м'язів у кров, аланін поглинається гепатоцитами і після перетворення в піруват використовується в глюконеогенезі
У печінці
Глюкоза
Піруват
Аланін
Кров
У м'язах
Глюкоза
Піруват
Аланін
16
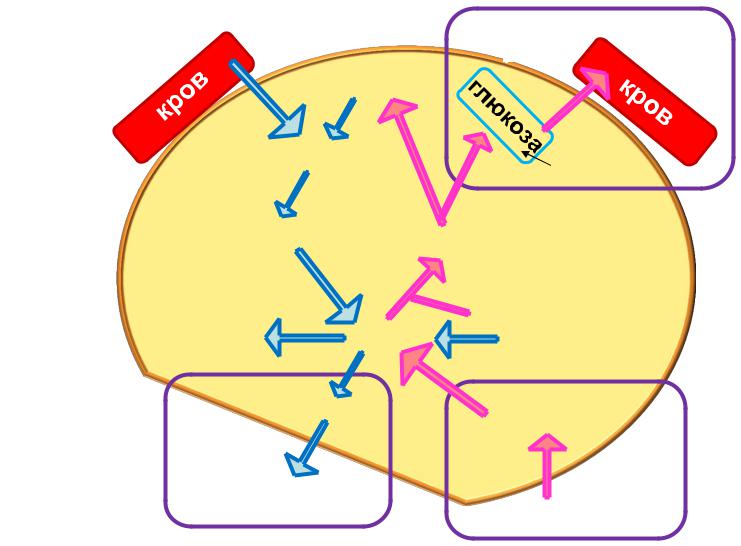
Кооперація між гліколізом і глюконеогенезом
У більшості |
У печінці, нирках |
|
|
клітин |
глікоген |
|
глюкоза |
ЕР |
|
глюкозо-6-Ф глюкозо-6-Ф
глюконеогенез
гліколіз
|
|
гліцерол |
СО2+Н2О |
ПВК |
амінокислоти |
лактат |
лактат |
|
|
Переважно у м'язах, |
|
еритроцитах |
У печінці, нирках |
|
ЕР – ендоплазматичний ретикулум
Гліколіз і глюконеогенез координуються за тканинною специфічністю з метою забезпечення залежних від глюкози енергетичних потреб,
необхідних для всіх клітин, що зустрічаються. |
17 |
|
Глюкагон
Інсулін
Вплив гормонів-антагоністів на обмін вуглеводів
Глюкагон
Гіпоглюкоземія, гіпоглікемія (зменшення рівня глюкоземії)
супроводжується збільшенням рівня секреції α-клітинами острівкової частини підшлункової залози глюкагону, який стимулює процеси глюконеогенезу та фосфороліз глікогену (за рахунок активації аденілатциклазної системи в мембранах гепатоцитів), і таким чином, підвищує рівень вільної глюкози.
Інсулін
Гіперглюкоземія, гіперглікемія (збільшення рівня глюкоземії)
супроводжується підвищенням рівня секреції β-клітинами острівкової частини підшлункової залози інсуліну, який підвищує ступінь проникності плазматичних мембран багатьох клітин для глюкози (окрім головного мозку). Інсулін зменшує швидкість ферментів глюконеогенезу в печінці, стимулює синтез ключових регуляторних ферментів гліколізу (посилюючи гліколітичний розпад глюкози) і синтетичні процеси утворення глікогену в печінці та м'язах, і таким чином, знижує рівень вільної глюкози.
18
Вплив гормонів-синергістів на обмін вуглеводів
Глюкагон, адреналін, глюкокортикоїди, соматотропін
стимулюють метаболічні процеси вуглеводів у бік підвищення рівня глюкози в крові
Адреналін (гормон мозкової частини наднирникових залоз) за
збільшення його секреції спостерігається підвищення вмісту глюкози у крові (гіперглюкоземія) за рахунок стимуляції фосфоролізу глікогену в м'язах і частково в печінці.
Глюкокортикоїди (наприклад, кортизол) підвищують рівень глюкози в
крові, наприклад, за стимуляції ФЕП-кінази (ензиму глюконеогенезу) та
ферментів, які перетворють деякі глюкогенні амінокислоти (серин, тирозин, триптофан) у субстрати глюконеогенезу.
Соматотропін (гормон аденогіпофіза) подібно до інсуліну збільшує
проникність плазматичних мембран клітин м'язової та жирової тканин для
глюкози, але на відміну від інсуліну активує глюконеогенез у печінці.
19
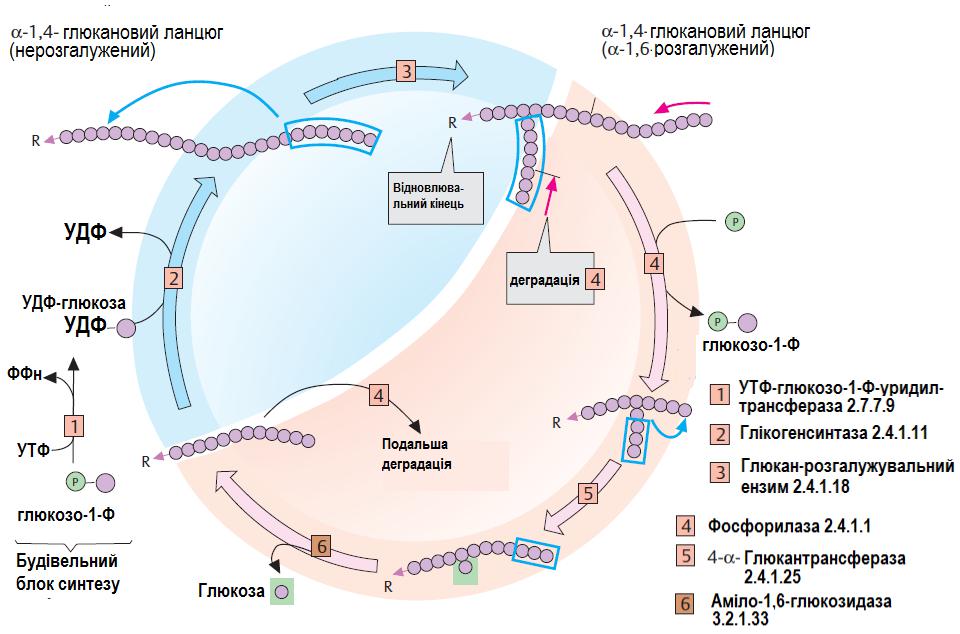
Метаболізм
глікогену
Синтез
Розпад
20