

Харченко Н. Н. Лесная зоология (лекции)
.pdf40
Класс II. Стеклянные губки (Hyalospongia)
Морские преимущественно глубоководные губки высотой до 50 см. Тело трубчатое, мешковидное, иногда в виде бокала. Почти исключительно
одиночные формы сиконоидного типа. Кремневые иглы, слагающие скелет, крайне разнообразны, в основе трехосные. Часто спаиваются концами, образуя решетки разной сложности. Характерная черта стеклянных губок — слабое развитие мезоглеи и слияние клеточных элементов в синцитиальные структуры. Типичный род Euplectella. У некоторых видов этого рода тело цилиндрическое, до 1 м в высоту, иглы у основания, втыкающиеся в грунт, достигают 3 м длины.
Класс III. Обыкновенные губки (Demospongia)
К этому классу принадлежит большинство современных губок. Скелет кремневый, спонгиновый или сочетание того и другого. Сюда относится
отряд четырехлучевых губок (Tetraxonia), скелет которых слагается четырехосными иглами с примесью одноосных. Характерные представители: шаровидные крупные геодии (Geodia), ярко окрашенные оранжево-красные морские апельсины (Tethya), комковидные яркие пробковые губки (сем. Suberitidae), сверлящие губки (сем. Clionidae) н многие другие. Второй отряд класса Demospongia — кремнероговые губки (Cornacuspongia). В состав скелета входят спонгин как единственный компонент скелета или в разных соотношениях с кремневыми иглами. Сюда принадлежат туалетные губки, немногочисленные представители пресноводных губок — бадяг из сем. Spongillidae, эндемичные байкальские губки сем. Lubomirskiidae.
Происхождение типа Spongia
В организации губок много признаков большой примитивности:
1.отсутствие настоящих дифференцированных тканей и органов,
2.чрезвычайная пластичность клеточных элементов.
3.отсутствие резко выраженной индивидуальности в колониях.
Все это свидетельство того, что губки — просто устроенные представители многоклеточных.
Если принять теорию Мечникова о происхождении многоклеточных, то легко видеть, что личинка, свойственная большинству губок,— паренхимула, по строению почти полностью соответствует гипотетической мечниковской фагоцителле. У нее имеется поверхностный, эктодермальный слой жгутиковых клеток и внутренний рыхлый слой клеток — энтодерма. Можно предположить,
41
что фагоцителла перешла к сидячему образу жизни и таким путем дала начало типу губок. При этом судьба клеточных слоев фагоцителлы у губок оказалась иной, чем у прочих многоклеточных («извращение» зародышевых листков): наружный эктодермальный слой жгутиковых клеток у губок дал начало пищеварительному слою хоаноцитов, который вместе с тем осуществляет кинетическую мерцательную водо-движущую функцию; внутренние энтодермальные клетки зародыша, которые у других групп животных дают начало энтодермальной кишке, у губок превращаются в клетки поверхности тела (дермальные) и в клеточные элементы мезоглеи. Все эти факты говорят о том, что отделение губок от ствола многоклеточных произошло очень рано, еще до того, как определилась окончательная судьба двух основных клеточных пластов тела.
Некоторые зоологи считают, что губки произошли от колониальных воротничковых жгутиконосцев независимо от прочих многоклеточных.
Другие полагают, что многоклеточные происходят общим стволом, от которого очень рано отделились губки.
Второй взгляд представляется более обоснованным потому, что личинка — паренхимула губок — сходна с планулой кишечнополостных. Это говорит об общности их происхождения.
Губки — очень древние организмы. Их ископаемые остатки многочисленны в кембрийских морских отложениях. Встречаются они и в протерозойских породах.
III. Надраздел Эуметазои.
К высшим многоклеточным относится множество типов и классов животных с разным уровнем организации. Но общими для них чертами являются: дифференцированность тканей и органов; присутствие нервных клеток; выраженная интегрированность и целостность отдельных особей; выражена радиальная или билатеральная симметрия тела. В соответствии с последним, эуметазои подразделяются на 2 раздела: Лучистые (радиальная симметрия во внешнем и внутреннем строении) и Билатеральные (двусторонняя симметрия).
Лучистые (Radiata). Характеризуются следующими основными особенностями организации: лучевой симметрией, двуслойностью строения, очень рано отделились губки.
Второй взгляд представляется более обоснованным потому, что личинка — паренхимула губок — сходна с планулой кишечнополостных. Это говорит об общности их происхождения.
42
Губки — очень древние организмы. Их ископаемые остатки многочисленны в кембрийских морских отложениях. Встречаются они и в протерозойских породах.
III. Надраздел Эуметазои.
К высшим многоклеточным относится множество типов и классов животных с разным уровнем организации. Но общими для них чертами являются: дифференцированность тканей и органов; присутствие нервных клеток; выраженная интегрированность и целостность отдельных особей; выражена радиальная или билатеральная симметрия тела. В соответствии с последним, эуметазои подразделяются на 2 раздела: Лучистые (радиальная симметрия во внешнем и внутреннем строении) и Билатеральные (двусторонняя симметрия).
Лучистые (Radiata). Характеризуются следующими основными особенностями организации: лучевой симметрией, двуслойностью строения, наличием кишечной (гастральной) полости и нервной системы диффузного типа.
Тело радиальных животных обладает гетерополярной осью, вокруг которой располагаются в радиальном порядке повторяющиеся морфологические структуры. Биологическое значение радиальной симметрии состоит в том, что такой организм имеет сходные пространственные возможности в захвате пищи и обороне от врагов.
Это преимущественно морские животные. Среди них выделяют 2 типа: Кишечнополостные (медузы, полипы со стрекательными клетками на щупальцах) и Гребневики (исключительно плавающие морские животные с рядами особых гребневидных пластинок, являющихся производными жгутиковых клеток).
Тип Кишечнополостные включает 3 класса: 1) Гидроидные, 2) Сцифоидные, 3) Коралловые полипы.
Это, главным образом, морские животные, реже пресноводные, ведущие сидячий или плавающий образ жизни. К ним относятся одиночные или колониальные полипы, а также медузы. Известно более 10 тыс. видов кишечнополостных. Они сочетают в себе признаки примитивной организации с чертами специализации к малоподвижному или неподвижному образу жизни. Специфичными признаками типа являются: наличие стрекательных клеток (орган защиты и нападения) и развитие с метаморфозом (двухслойная личинкапланула), реже развитие прямое. Все имеют гастральный тип строения (двухслойный мешок с полостью внутри). Но в связи с разным образом жизни
43
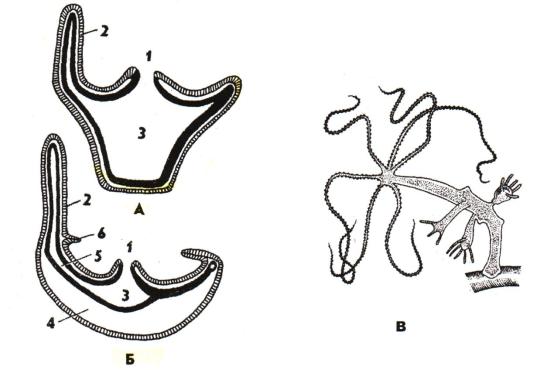
существуют в двух морфо-экологических формах - полип и медуза (рис. 26)
Рис. 26. Строение гидроидного полипа, гидроидной медузы (по Холодковскому) и пресноводной гиды Hydra olidactis с почками (по Полянскому): А - полип, Б - медуза (продольный разрез): 1 - рот, 2 - щупальце, 3 - гастральная полость, 4 - мезоглея, 5 - радиальный канал, 6 – парус, В - пресноводная гидра
Это разные жизненные формы: медуза - планктонная, а полип - бентосная прикрепленная. В ряде случаев кишечнополостные образуют колонии из полипоидных и медузойдных особей.
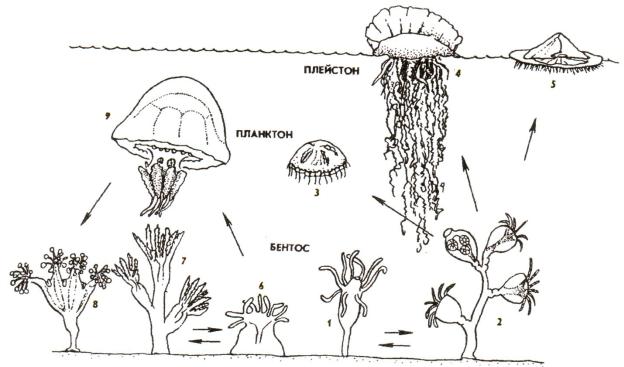
От исходной для кишечнополостных жизненной формы бентобионтов полипоидных, ведущих прикрепленный образ жизни, прослеживаются переходы к колониальным полипам, а также к плавающим медузам (рис. 27).
44
Рис. 27. Экологическая радиация кишечнополостных: 1 - гидроидный полип, 2 - морской колониальный гидроидный полип, 3 - гидромедуза, 4 - сифонофора, 5 - плавающий полип, 6 - коралловый полип (одиночный), 7 - колониальный коралловый полип, 8 - сидячая медуза, 9 - сцифоидная медуза (по Шаровой)
1. Класс Гидрозои (Hydrozoa.)
Чаще морские, реже пресноводные. Жизненная форма - одиночный полип. Морские гидроидные полипы преимущественно колониальны, в цикле развития присутствует гидромедуза - половая стадия.
2. Класс Сцифоидные медузы (Scyphozoa)
Свободноживущие обитатели морей. Передвигаются по реактивному принципу. В цикле развития присутствует недолговечная стадия полипа, дающая начало новым медузам путем почкования.
3. Класс Коралловые полипы (Anthozoa)
Прикрепленный образ жизни в морских водах. Обычно колониальные, есть и одиночные (актинии). В жизненном цикле отсутствует стадия медузы и смена поколений.
По сравнению с гидрозоями имеют эктодермальную глотку, гастральная полость подразделяется септами на камеры.
У большинства имеется эктодермальный скелет (известковый, кремниевый).
45
ЖИЗНЕННЫЙ ЦИКЛ КИШЕЧНОПОЛОСТНЫХ, ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Из современных нам кишечнополостных животных наиболее просто завершается жизненный цикл коралловых полипов. Оплодотворенное яйцо начинает дробиться. Сначала оно делится надвое, затем каждая из образовавшихся клеточек в свою очередь также разделяется, и так далее. В результате образуется большое количество мелких клеток, расположенных в один слой и имеющих вид маленького полого шарика. Вслед за этим часть клеток начинает погружаться внутрь, в результате чего получается двуслойный зародыш. Из его внутреннего слоя впоследствии формируется энтодерма, а из наружного — эктодерма будущего полипа. Эктодерма покрыта многочисленными мелкими ресничками, при помощи которых зародыш получает способность плавать; с этого момента он превращается в личинку, называемую планулой. Планула не способна питаться и размножаться. Она некоторое время плавает в толще воды, затем садится на дно, прикрепляясь к нему передним концом. Вскоре после этого на заднем (теперь уже верхнем) конце планулы прорывается ротовое отверстие и образуется венчик щупалец. Так возникает первый полип. У колониальных форм этот полип вскоре выпочковывает на себе других полипов, те в свою очередь следующих и т. д. Возникает колония. По достижении колонией определенной степени развития составляющие ее полипы начинают размножаться также и половым путем, образуя яйца. На этом цикл замыкается. Иначе протекает жизненный цикл у гидроидных и сцифоидных. Дробление яйца и развитие планулы происходит у них так же, как у коралловых полипов, аналогичным способом возникает и первый полип — основатель колонии, а потом и вся колония (у сцифоидных колонии обычно не образуется). Однако полипы гидроидных и сцифоидных совершенно лишены способности к половому размножению, зато они выпочковывают или отделяют не только подобных себе полипов, но совершенно не похожих на них медуз. Медузы и являются половыми особями. Образование медуз у гидроидных и сцифоидных происходит по-разному. У гидрополипов медузы возникают из почек. Вначале такая почка внешне похожа на обычную почку, из которой развивается полип, но затем появляются отличия. Медузоидная почка сильно увеличивается в размере, становится полупрозрачной, внутри ее появляются 4 радиально расположенных канала и отходящий от места их скрещивания ротовой хоботок. Теперь молодая медузка внешне напоминает маленький колокольчик или зонтик. Вскоре она отрывается от колонии полипов и уплывает. Медузы самостоятельно питаются и
46
увеличиваются в размерах, затем у них возникают половые железы. Яйца и сперма выбрасываются прямо в морскую воду, где происходит оплодотворение яиц. Из яиц возникает личинка — планула. У сцифоидных, как правило, полипы (их называют сцифистомами) не образуют колоний: возникшие в результате почкования полипчики отделяются от материнской сцифистомы и ведут самостоятельный образ жизни. Вскоре каждый такой полип претерпевает серьезные изменения: его щупальца укорачиваются, а на теле появляется ряд кольцевидных перетяжек, которые становятся все более и более глубокими. Затем начиная с верхнего конца от полипа, отделяются дисковидные медузы (на этой стадии их называют эфирами), которые переворачиваются ротовой стороной вниз и уплывают. Эфиры питаются, растут и превращаются в крупных половозрелых сцифоидных медуз, размножающихся исключительно половым путем. Таким образом, на примере гидроидных и сцифоидных мы имеем возможность познакомиться с весьма своеобразным явлением, называемым чередованием поколений. Действительно, в жизненном цикле этих двух групп кишечнополостных наблюдается правильная смена донного, сидячего, часто колониального полипоидного поколения, размножающегося вегетативно, с поколением свободноплавающим, одиночным, медузоидным, размножающимся половым путем. Разные поколения ведут различный образ жизни и устроены неодинаково, поэтому до открытия явления чередования поколений их считали самостоятельными группами животных. Чередование поколений имеет очень большое значение для жизни гидроидных и сцифоидных: прикрепленный образ жизни полипов препятствует их расселению, а медузы, покинув колонию, могут далеко уноситься морскими течениями и разбрасывать яйца на очень большом пространстве, заселяя все новые и новые области. Чередование различно устроенных поколений довольно широко встречается в животном мире. Кроме кишечнополостных, оно обнаружено у некоторых плоских, круглых и кольчатых червей, у ряда ракообразных и даже у низших хордовых. Именно у представителей хордовых — сальп — известный зоолог и поэт Шамиссов 1819 г. открыл чередование поколений. Это открытие повлекло за собой поиски новых случаев чередования поколений. В 1841 г. М. Саре описал чередование поколений у двух сцифоидных медуз— аурелии (Aurelia) и цианеи (Cyanea), а годом позже Стенструп установил, что гидроидные также развиваются с чередованием поколений. Исследования Шамиссо, Сарса и Стенструпа явились значительным событием для зоологов прошлого века, благодаря им пришлось пересмотреть многие старые представления о биологии и систематике. Описанный выше способ чередования поколений у кишечнополостных называют метагенезом, а животных с этим типом жизненного цикла — мета —
47
генетическими. Для метагенеза характерно правильное чередование полового поколения с поколением, размножающимся вегетативно, путем деления или почкования. Не следует думать, что жизненный цикл у всех видов гидроидных
исцифоидных идет строго по той схеме, с которой мы уже ознакомились. В большинстве случаев от такого типичного проявления метагенеза имеются более или менее заметные отклонения, зависящие от целого ряда причин. Главной причиной, вызывающей нарушение правильного метагенеза, следует считать различие условий среды, в которых обитают полипы и медузы. В самом деле, придонный, неподвижный образ жизни полипов неизбежно вызывает у них появление целого ряда приспособлений, отвечающих условиям их существования. Так как полипы неподвижно сидят на дне, они не нуждаются в особых органах чувств, позволяющих им на расстоянии узнавать о приближении врага или добычи: ни приблизиться к добыче, ни уйти от врагов они все равно не могут. Поэтому полипы сохранили одно только чувство осязания, позволяющее им ощупью ловить добычу, прикоснувшуюся к расставленным щупальцам. Другой способ защиты полипов заключается в образовании наружного скелета. Чем прочнее и тяжелее скелет, тем надежнее он защищает полипов и лучше удерживает всю колонию на морском дне. Указанные выше особенности полипов характерны не для одних только кишечнополостных — они вырабатываются у многих прикрепленных водных организмов. Общим свойством большинства сидячих животных является также образование колоний. Очень характерно оно и для полипов кишечнополостных
идля сифонофор. С образованием колоний часто связано разделение функций ее членами. Особенно ярко это проявляется у гидроидных полипов. Только у очень немногих видов все полипы устроены одинаково и каждый из них может отпочковывать медуз. Гораздо чаще одни полипы, снабженные щупальцами и ртом, ловят для колонии добычу, но не способны размножаться, тогда, как другие лишены щупалец и рта, зато приспособлены исключительно к почкованию медуз. Так дело обстоит у широко распространенного гидроида обелии (Obelia geniculata). У таких видов правильность метагенеза нарушается, так как часть полипов (кормящие особи) не дает полового потомства. Чем сложнее становится колония, тем большее значение приобретает она в жизненном цикле. Роль медуз при этом уменьшается. Большая колония способна прокормить не только полипов, но и медуз. Поэтому медузы долгое время не отрываются от колонии и покидают ее только тогда, когда их половые продукты созревают. Наконец, имеется много таких видов гидроидных, колония которых настолько усложнена, что медузы вообще прекращают отрываться, и становятся такими же прикрепленными организмами, как
48
полипы. При этом характерные признаки медузы — зонтик, глазки, каналы гастроваскулярной системы — постепенно утрачиваются, и медуза становится похожей на набитый яйцами мешочек. У пресноводных гидр медузы вовсе не образуются. Но вернемся к типичному метагенезу и проследим за свободноплавающими медузами. Их образ жизни резко отличается от того, который ведут полипы. Медузы— подвижные животные, и потому у них вырабатываются приспособления для движения и ориентировки в пространстве. В первую очередь это касается развития зонтика и мускулатуры, а также органов чувств. У отдельных видов эти приспособления развиты настолько хорошо, что медузы могут далеко уплывать от тех мест, где они начали свою самостоятельную жизнь, отделившись от полипа. Часто они попадают в такие места, где нет подходящих условий для развития полипоидного поколения, например в открытое море, над большими глубинами. Планулы таких медуз, не найдя подходящего места для прикрепления, погибают. Но так обстоит дело далеко не со всеми видами гидроидных и сцифоидных медуз. Часть из них приспособилась к жизни именно в поверхностных слоях воды в открытом море, но при этом изменился и их жизненный цикл. У гидромедузы бугенвиллии (Bougainvillia platygaster) яйца не выбрасываются в воду, а развиваются в половой железе самки, здесь же происходит и образование маленьких полипчиков, которые выпочковывают новых медуз. Случай этот не единственный, развитие полипов прямо на половой железе медузы характерно также и для одного вида кампанулярии (Campanularia maccrady), и для некоторых других медуз. У этих видов полипоидное поколение хотя и имеется, но оно перестало быть донным и не образует больших колоний. У некоторых видов подавление полипоидной стадии пошло еще дальше, и полипы вовсе исчезли из жизненного цикла. Из планулы таких медуз возникают не полипы, а такие же медузы. Так протекает жизненный цикл большой группы гидроидных медуз—трахилид, а из сцифоидных у видов рода пелагия (Pelagia), живущих в открытом море. Как видно из приведенных примеров, метагенетический жизненный цикл весьма пластичен. Пока он является выгодным для процветания вида, он сохраняется, но как только условия меняются и одно из поколений получает некоторое преимущество, другое поколение подавляется и может совсем исчезнуть.
49
СТРЕКАТЕЛЬНЫЕ КЛЕТКИ КИШЕЧНОПОЛОСТНЫХ И ДЕЙСТВИЕ ИХ ЯДА
Характернейшей особенностью всех кишечнополостных является наличие у них особых стрекательных, или крапивных, клеток. Эти клетки развиваются из недифференцированных промежуточных клеток эктодермы. Каждая стрекательная клетка содержит хитиноидную капсулу овальной или продолговатой формы. Стенка наружного конца капсулы впячивается внутрь ее и имеет вид тонкой спирально закрученной трубочки, называемой стрекательной нитью. Полость капсулы заполнена ядовитой жидкостью.
На внешней поверхности стрекательной клетки имеется чувствительный волосок — книдоциль. Прикосновение к волоску вызывает моментальное раздражение клетки, проявляющееся в стремительном, подобном выстрелу выворачивании стрекательной нити. В зависимости от деталей строения и способа воздействия на врага или жертву различают несколько типов стрекательных капсул. Часть стрекательных капсул имеет длинную, снабженную шипами нить. При «выстреле» такой капсулы нить вонзается в тело жертвы, а через ее полость изливается содержимое капсулы, вызывающее общее или местное отравление. У других капсул нить короткая, лишенная шипов. Такие нити только опутывают жертву. Наконец, имеются липкие нити, которые удерживают добычу приклеиванием. Они же могут служить и для временного прикрепления самих носителей стрекательных клеток при передвижении.
Стрекательные клетки располагаются по всей поверхности тела кишечнополостных, но их наибольшие скопления наблюдаются на щупальцах и вокруг ротового отверстия, т. е. в местах, где они нужнее всего. После «выстрела» стрекательная клетка погибает и на ее месте или поблизости от него развивается новая стрекательная клетка.
Весьма интересно, что стрекательные капсулы могут действовать и после гибели животного. Так, прикосновение к мертвым, выброшенным на берег медузам может вызвать довольно сильные покраснения кожи, сопровождающиеся ощущением жжения.
Стрекательные клетки, особенно содержащие ядовитую жидкость в капсулах, являются грозным оружием, которое используется кишечнополостными и для защиты и для нападения. Несомненно, что процветание этих низко организованных животных в значительной мере объясняется наличием у них стрекательных клеток. Мелкие животные,