

1.2 Биохимические процессы, протекающие при сбраживании спиртового сусла
Спирт образуется в результате сбраживания сахаров дрожжевыми клетками под действием ферментов. Под брожением следует понимать такие энергетические процессы, в которых органические углеродсодержащие соединения функционируют одновременно и как доноры, и как акцепторы электронов. При брожении роль конечного окислителя, или акцептора электронов, играет обычно какая-нибудь органическая молекула, которая образуется в ходе самого брожения.
Брожение – это внутренний окислительно-восстановительный процесс, называемый также анаэробным дыханием в отличие от аэробного дыхания, когда конечным акцептором электрона является кислород.
Из смеси сахаров, содержащейся в сусле, быстрее всех сбраживаются фруктоза и глюкоза. Ди- и трисахариды сусла, которые непосредственно не сбраживаются дрожжами, должны предварительно подвергнуться гидролизу соответствующими ферментами дрожжевых клеток. Из дисахаридов наиболее легко и быстро сбраживается сахароза, которая гидролизуется (инвертируется) уже в начале брожения. Возможно, это объясняется тем, что фермент инвертаза (сахараза) локализована в поверхностном слое дрожжевой клетки. При переходе дрожжей от анаэробных условий к аэробным ослабляется способность их сбраживать глюкозу и мальтозу, а сахаразная активность повышается в 2,5 раза. Мальтозу дрожжи начинают потреблять только тогда, когда фруктозы и глюкозы в сусле почти не остается. Трисахарид рафинозу, сбраживают полностью или на одну треть, это зависит от расы дрожжей.
Для сбраживания сусла в него вводят определенное количество заранее размноженных дрожжей. Дрожжевая масса, вводимая в сусло, состоит из большого числа отдельных клеток. Несмотря на незначительную величину дрожжевой клетки, общая поверхность дрожжевой массы очень большая, так как только в 1 г прессованных дрожжей находится примерно 14 млрд. клеток. При эквивалентном среднем диаметре клетки 8 мкм площадь ее поверхности будет равна 5 ·10-5мм2. В соответствии с этим 10 г отпрессованных сухих дрожжей с 25 % сухих веществ имеют поверхность площадью от 9 до 10 м2 . При такой сильно развитой поверхности адсорбция составных веществ сусла происходит довольно быстро.
Брожение всегда включает промежуточные окислительно-восстановительные реакции, но никогда не приводит к полному окислению органических веществ. При спиртовом брожении распад сахаров заканчивается образованием этилового спирта и диоксида углерода. Следует также отметить, что одновременно осуществляются и биосинтетические реакции различных побочных продуктов, составляющих примеси этилового спирта.
Основное значение брожения состоит в перестройке структуры молекул гексозы или другого углеродсодержащего соединения. Эти перестройки обусловливают биохимические превращения исходного субстрата, протекание окислительно-восстановительных процессов и образование в конечном итоге высокоактивного и лабильного в химическом отношении метаболита, которым является пировиноградная кислота.
Спиртовое брожение осуществляется ферментативным путем, при котором не участвует кислород. Однако брожение включает в себя окислительно-восстановительные реакции. Об этом особенно отчетливо свидетельствуют конечные продукты спиртового брожения: этанол можно рассматривать как относительно восстановительное соединение, поскольку его молекула содержит определенное количество водорода, а С02 – как относительно окислительное, поскольку в его молекуле водород отсутствует.
1.2.1 Химизм спиртового брожения
Процессы спиртового брожения сопровождаются образованием АТФ из АДФ и фосфата. Схема спиртового брожения показана на рисунке 1.
Без одновременного фосфорилирования АДФ они вообще не могут протекать. Суммарное уравнение спиртового брожения имеет следующий вид:
С6Н12О6 + 2ФН + 2АДФ = 2СН3СН2ОН + 2СО3 + 2АТФ + 2Н2О,
причем оно включает ряд стадий спиртового брожения.
На первом этапе различные гексозы вовлекаются в процесс диссимиляции, фосфорилируясь за счет АТФ и образуя общий продукт — глицеральдегид-3-фосфат. Второй этап представляет собой процесс, общий для всех сахаров. Он включает и окислительно-восстановительные реакции, и механизмы накопления энергии – стадии, в ходе которых АДФ фосфорилируется до АТФ.
Первая стадия спиртового брожения начинается с фосфорилирования D-глюкозы за счет АТФ:
Глюкоза + АТФ → Глюкозо-6-фосфат + АДФ
Реакция фосфорилирования как бы запускает процесс брожения. Нейтральная молекула D-глюкозы при этом активируется для участия в последующих этапах диссимиляции. В результате фосфорилирования за счет АТФ глюкоза превращается в отрицательно заряженную частицу – глюкозо-6-фосфат. Этот процесс катализируется ферментами двух типов: гексокиназой и глюкокиназой, которые различаются между собой по специфичности действия в отношении сахаров. Превращение глюкозо-6-фосфата во фруктозо-6-фосфат происходит в результате реакции изомеризации.
Данный процесс катализирует фермент фосфоглюкоизомераза. При фосфорилировании D-фруктозо-6-фосфата с образованием фруктозо-1,6-дифосфата используется еще одна молекула АТФ при участии фермента фосфофруктокиназы.
Фруктозо-6-фосфат + АТФ → Фруктозо-1,6-дифосфат
Роль специфического акцептора фосфата в реакции играет фруктозо-6-фосфат. Роль донора фосфата могут выполнять помимо АТФ также УТФ и ИТФ (уридинтрифосфат и инозинтрифосфат).
Фосфорилирование фруктозо-6-фосфата – важный этап, при помощи которого осуществляется регуляция брожения. Фосфофруктокиназа относится к числу аллостерических, или регуляторных, ферментов. Поскольку фосфофруктокиназа ингибируется АТФ и цитратом (в высоких концентрациях) и стимулируется АДФ и АМФ, считается, что она представляет собой поливалентный аллостерический фермент, т. е. подчиняется действию более чем одного специфического модулятора (метаболита-ингибитора).
Фосфофруктокиназа, как регуляторный фермент, катализирует реакцию, лимитирующую общую скорость спиртового брожения.
Расщепление фруктозе-1,6-дифосфата на глицеральдегид-3-фосфат и диоксиацетонфосфат осуществляется альдолазой. Реакция представляет собой обратимую альдольную конденсацию:
D-Фруктозо-1,6-дифосфат→Диоксиацетонфосфат+D-Глицеральдегид-З-фосфат
Свободная энергия в этой реакции выражается довольно большой положительной величиной. В реакциях этого типа, в которых одна молекула исходного вещества расщепляется с образованием двух молекул продукта, на положение равновесия сильно влияет концентрация исходного вещества. Чем меньше начальная концентрация фруктозо-1,6-дифосфата, тем большая доля его должна расщепляться для того, чтобы было достигнуто состояние равновесия.
Глюкоза
АТФ; Мg2+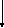

Гексокиназа
Глюкозо-6-фосфат + АДФ – ~Р
Глюкозофосфатизомераза

Фруктазо-6-фосфат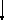
Фосфофруктокиназа
Фруктозо-1,6-дифосфат
+ АДФ – ~Р Глицерин
Альдолаза
(Zn2+,Co2+,Ca2+)
Фосфодиоксиацетон
Изомераза 3-Фосфоглицериновый 
альдегид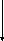


Н3РО4
неорганическя
1,3-Дифосфоглицериновая кислота
Триозофосфадегидрогеназа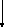

2НАД 2НАД Н2





1,3-Дифосфоглицериновая
кислота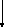


АДФ Фосфотрансфераза

АТФ

3-Фосфоглицериновая
кислота
Фосфоглицератмутаза
2-Фосфоглицериновая
кислота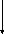
Н2О Энолаза

Mg2+,Mn2+,Zn2+
Фосфоэнолпировиноградная
кислота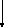
АДФ Фосфотрансфераза


АТФ К+
Энолпировиноградная
кислота
Пировиноградная
кислоа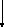
Карбоксилаза

Уксусный альдегид
+ СО2

Алкогольдегидрогеназа
Этиловый спирт
Рисунок 1 – Схема спиртового брожения
Молекула альдолазы (молекулярная масса 150000) состоит из четырех субъединиц. В кислой среде фермент диссоциирует на неактивные субъединицы; в нейтральной среде субъединицы легко и самопроизвольно воссоединяются, образуя активный фермент. Альдолаза содержит свободные SH-группы; некоторые из них играют важную роль в ее каталитической активности. Фермент проявляет строгую специфичность в отношении диоксиацетонфосфата, но вместо глицеральдегид-3-фосфата может катализировать реакцию с другими альдегидами, например с ацетальдегидом или незамещенным глицериновым альдегидом.
Полагают, что альдолаза образует фермент-субстратный комплекс с диоксиацетонфосфатом и другими кетозо-1-фосфатами.
Таким образом, перечисленная цепь реакций завершают первую стадию спиртового брожения, в которой молекула углевода путем двух фосфорилирований и расщепления подготавливается для второй стадии.
На второй стадии протекают окислительно-восстановительные реакции и реакции фосфорилирования, в процессе которых генерируется АТФ. Поскольку из одной молекулы глюкозы образуются 2 молекулы глицеральдегид-3-фосфата, в последующих реакциях обе половины молекулы глюкозы претерпевают одинаковые превращения.
При окисление глицеральдегид-3-фосфата до 1,3-дифосфоглицерата энергия, освобождающаяся при окислении альдегидной группы глицеральдегид-3-фосфата, сохраняется в форме высокоэнергетического продукта окисления 1,3-дифосфоглицерата.
В этой реакции проявляется одновременно ферментативный и химический механизм, с помощью которого энергия, высвобождающаяся в ходе окисления органических молекул, может запасаться в молекулах АТФ. Катализирует эту реакцию глицеральдегид-3-фосфат-дегидрогеназа. Фермент получен в кристаллическом виде (молекулярная масса 140 000) из дрожжей и мышц животных. Молекула его содержит 4 идентичные субъединицы, каждая из которых представляет собой одиночную полипептидную цепь, состоящую примерно из 330 аминокислотных остатков.
Ниже представлена суммарная реакция, катализируемая глицеральдегид-3 -фосфат-дегидрогеназой:
D-Глицералъдегид-З-фосфат + НАД+ + ФН → 1,3-дифосфоглицерат + НАДН + Н+
В этой реакции к окислителю НАД+ присоединяется электрон от альдегидной группы D-глицеральдегид-З-фосфата. Известно, что кофермент НАД является компонентом козимазы. Он служит переносчиком электронов от D-глицеральдегид-3-фосфата, играющего роль донора электронов, к пирувату, образующемуся на одном из более поздних этапов брожения.
Следовательно, окисление за счет НАД+ представляет собой экзергонический процесс, который в обычных условиях идет в сторону завершения, тогда как эндергоническая реакция образования 1,3-дифосфатглицерата в таких условиях идти не может. В суммарной ферментативной реакции эндергонический процесс сопряжен с экзергоническим: энергия, высвобождающая при окислении альдегида, запасается в высокоэнергетической фосфатной группе 1,3-дифосфоглицерата.
Образовавшийся 1,3-дифосфоглицерат вступает далее в ферментативную реакцию с АДФ, катализируемую фосфоглицераткиназой. При этом фосфатная группа переносится на АДФ с образованием АТФ и 3-фосфоглицерата:
1,3-Дифосфоглицерат + АДФ → 3-Фосфоглицерат + АТФ
Реакция эта сильно экзергоническая, позволяющая предыдущей реакции идти до полного завершения.
Последующее превращение 3-фосфоглицерата в 2-фосфоглицерат осуществляется посредством фермента фосфоглицеромутазы. Фосфатная группа переносится из положения 3 в глицериновой кислоте в положение 2:
3-Фосфоглицерат → 2-фосфоглицерат
Фермент фосфоглицеромутаза существует в двух формах: 1) для одной из них в качестве промежуточного продукта необходим 2,3-дифосфоглицерат, 2) «собственно» фосфоглюкомутаза, механизм действия которой является аналогом для первой.
При дегидратация 2-фосфоглицерата с образованием фосфоенолпирувата, во второй реакции спиртового брожения образуется высокоэнергетическая связь; реакция катализируется енолазой:
2-фосфоглицерат → Фосфоенолпируват + Н2О
Рассматриваемая реакция формально представляет собой отщепление молекулы Н2О от 2-го и 3-го углеродных атомов 2-фосфоглицерата, одновременно ее можно также рассматривать как внутримолекулярный окислительно-восстановительный процесс; так, при отщеплении молекулы Н2О степень окисления 2-го углеродного атома увеличивается, а 3-го уменьшается. Гидролитическое отщепление фосфатной группы от исходного вещества (2-фосфоглицерата) и от продукта (фосфоенолпирувата) сопровождается совершенно различным изменением стандартной свободной энергии; для фосфоглицерата оно равно приблизительно – 17,6 кДж, а для фосфоенолпирувата – 62,16 кДж. Тем самым дегидратация молекулы 2-фосфоглицерата, в результате которой образуется фосфоенолпируват, сопровождается значительным перераспределением энергии внутри молекулы.
Следующая реакция переноса фосфатной группы от фосфоенолпирувата на АДФ с образованием пирувата и АТФ катализируется пируваткиназой:
Фосфоенолпируват + АДФ → Пируват + АТФ
При декарбоксилирования пирувата образуется ацетальдегид:
Пируват → Ацеталъдегид + СО2
Катализирует эту реакцию пируватдекарбоксилаза, причем она характеризуется полной необратимостью. Фермент требует наличия ионов Mg2+ и кофермента тиаминпирофосфата (ТПФ, или кокарбоксилаза), который представляет собой эфир пирофосфорной кислоты и тиамина, необходимого многим микроорганизмам в качестве фактора роста и в качестве витамина В1.
Заключительная реакция спиртового брожения обеспечивает восстановление ацетальдегида до этанола под действием фермента алкогольдегидрогеназы. В качестве восстановителя здесь выступает НАДН+Н+:
Ацеталъдегид + НАДН +Н+ → Этанол + НАД+
Суммарное уравнение спиртового брожения можно поэтому написать в следующем развернутом виде:
Глюкоза + 2АТФ + 2НАД+ + 2ФН+ 4АДФ + 2НАДН + 2Н+ → 2-Этанол + 2 АДФ + 2НАДН + 2Н+ + 2НАД + 4АТФ + 2Н2О
Таким образом, процесс спиртового брожения состоит в превращении одной молекулы D-глюкозы в две молекулы этанола и двух молекул АДФ в две молекулы АТФ; кроме того, от глицеральдегид-3-фосфата к пирувату переносится четыре электрона в форме 2НАДН+2Н+. Реакции, обеспечивающие выделение энергии, сохраняют ее в форме АТФ. Причем, спиртовое брожение осуществляется с помощью 12 ферментов, катализирующих этот процесс.