

Biologia_V_N_Yarygin_Tom_1_2008
.pdfобеспечивают эффективную рекомбинацию аллелей и групп сцепления генов в гаметах, образуемых одним организмом.
Оплодотворение. Случайная встреча разных гамет при оплодотворении приводит к тому, что среди особей вида практически невозможно появление двух генотипически одинаковых организмов. Достигаемое с помощью описанных процессов генотипическое разнообразие особей предполагает наследственные различия между ними на базе общего видового генома.
Таким образом, геном как высший уровень организации наследственного материала благодаря мейозу и оплодотворению сохраняет свои видовые характеристики. Но одновременно эти же процессы обеспечивают индивидуальные наследственные различия особей, в основе которых лежит рекомбинация генов и хромосом, т.е. комбинативную изменчивость. Комбинативная изменчивость, проявляющаяся в генотипическом разнообразии особей, повышает выживаемость вида в изменяющихся условиях его существования.
3.6.2.4. Изменения геномной организации наследственного материала. Геномные мутации
Рассмотренные выше механизмы рекомбинации наследственного материала (кроссинговер, расхождение гомологичных хромосом и независимое поведение негомологичных хромосом в анафазе I мейоза, оплодотворение) при закономерном их течении обусловливают комбинативную изменчивость, но не изменяют общей структуры генома как видовой характеристики. Эволюционно сложившаяся у данного вида сбалансированность по дозам отдельных генов, распределение этих генов по группам сцепления остаются стабильной характеристикой генома каждого вида. Однако как на генном и хромосомном уровнях организации наследственного материала, так и на геномном уровне он способен приобретать мутационные изменения. Эти изменения могут использоваться как эволюционный материал. При этом ускоренные темпы эволюционного процесса, наблюдаемые на отдельных этапах исторического развития, как правило, бывают обусловлены не столько накоплением генных мутаций, сколько существенными изменениями структуры именно всего генома. К последним относятся изменения дозового соотношения различных генов и изменение состава групп сцепления внутри генома.
Причиной структурных изменений генома может быть нарушение тех процессов, которые в норме обеспечивают его устойчивость, в первую очередь процессов, протекающих в мейозе.
Так, нарушение кроссинговера, приводящее к обмену неравноценными участками ДНК между хроматидами, может привести к утрате или удвоению определенной нуклеотидной последовательности в них. Если это затрагивает структуру отдельного гена, то возможно возникновение генной мутации с изменением количества нуклеотидов в нем (см. разд. 3.4.2.3). Если при неравноценном обмене затронут участок хроматиды, содержащий несколько генов, изменяется доза этих генов в геноме. Он либо лишается каких-то генов (деления),
161
либо эти гены оказываются в геноме в двойном количестве (дупликапия). Изменение дозового соотношения отдельных генов наблюдается также при разных видах хромосомных перестроек, не обязательно связанных с неравноценным кроссинговером (см. разд. 3.5.3.3).
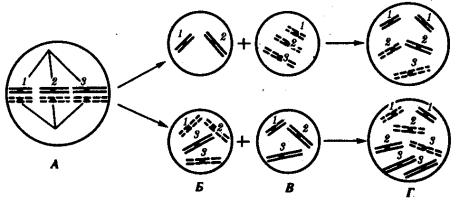
Нарушение расхождения бивалентов в анафазе I мейоза является причиной изменения количества хромосом в гаплоидном наборе гамет. Нерасхождение отдельного бивалента приводит к появлению одной гаметы, лишенной данной хромосомы, и другой, имеющей эту группу сцепления в двойном количестве (рис. 3.76). Оплодотворение таких гамет нормальными половыми клетками приводит к появлению особей, в кариотипе которых изменено общее число хромосом за счет уменьшения (моносомия) или увеличения (трисомия) числа отдельных хромосом. Нарушения структуры генома, заключающиеся в изменении количества отдельных хромосом, называют анэуплоидией.
Рис. 3.76. Нарушение расхождения отдельных бивалентов (1, 2, 3) в мейозе как причина возникновения анэуплоидий:
А — метафаза 1 мейоза; Б — образование аномальных гамет в результате нарушения расхождения 3-го бивалента в анафазе I мейоза; В — оплодотворение аномальных гамет нормальными гаметами другого пола; Г — образование зигот с анэуплоидным кариотипом (моносомия или трисомия по 3-й хромосоме, соответственно сверху и снизу)
В том случае, если в целом повреждается механизм распределения гомологичных хромосом между полюсами веретена (что наблюдается при его разрушении), клетка остается неразделившейся. Во второе деление мейоза она вступает не гаплоидной, а диплоидной. Из нее образуются диплоидные гаметы. Оплодотворение таких гамет приводит к образованию триплоидных организмов. Увеличение в кариотипе зиготы числа наборов хромосом называют полиплоидией.
Такие структурные изменения наследственного материала довольно часто встречаются в природе у растений, что обеспечивает у них относительно быстрые темпы видообразования. Полиплоидизацию путем искусственного разрушения
162
веретена деления с помощью колхицина широко применяют в селекции при выведении новых сортов растений.
Структурные изменения генома могут выражаться в ином распределении генов по группам сцепления. Когда отдельные хромосомы соединяются по типу робертсоновской транслокации или, наоборот, из одной хромосомы образуются две самостоятельные, это ведет к изменению числа групп сцепления в геноме (см. разд. 3.5.3.3). При реципрокных транслокациях между негомологичными хромосомами или при инверсиях изменяется место положения отдельных генов, что нередко сказывается на характере их функционирования (эффект положения).
Любые мутационные изменения в наследственном материале гамет — генеративные мутации —становятся достоянием следующего поколения, если такие гаметы участвуют в оплодотворении. Поэтому отклонения в течении митоза или мейоза в клетках-предшественницах гамет имеют большое эволюционное значение. Если же мутации любого ранга (генные, хромосомные или геномные) возникают в соматических клетках — соматические мутации — они передаются только потомкам этих клеток, т.е. не выходят за пределы данного организма. Исключение составляют соматические мутации, возникшие в клетках органов вегетативного размножения, от которых они передаются новому поколению организмов. Одной из причин соматических мутаций являются патологические митозы. При нарушении нормального течения митоза (нерасхождение хроматид отдельных хромосом, многополюсные митозы и т.д.) дочерние клетки получают аномальную наследственную программу и их дальнейшее развитие отклоняется от нормы. Патологические митозы часто наблюдаются в клетках злокачественных опухолей.
Таким образом, несмотря на существование механизмов, обеспечивающих стабильность структуры генома, на этом уровне организации наследственного материала могут появляться эволюционно значимые изменения. Они способны обеспечить достаточно резкий скачок в ходе исторического развития живой природы.
3.6.3.Особенности организации наследственного материала
упро- и эукариот
Геном современных прокариотических клеток характеризуется относительно небольшими размерами. У кишечной палочки (Е. coli) он представлен кольцевой молекулой ДНК длиной около 1 мм, которая содержит 4·106 пар нуклеотидов, образующих около 4000 генов. Основная масса ДНК прокариот (около 95%) активно транскрибируется в каждый данный момент времени. Как было сказано выше, геном прокариотической клетки организован в виде нуклеоида — комплекса ДНК с негистоновыми белками (см. разд. 3.5.2.4).
У эукариот объем наследственного материала значительно больше. У дрожжей он составляет 2,3 · 107 п.н., у человека общая длина ДНК в диплоидном хромосомном наборе клеток — около 174 см. Его геном содержит 3·109 п.н. и включает по последним данным 30—40 тыс. генов.
163
У некоторых амфибий и растений геном характеризуется еще большими размерами, достигающими 1010 и 1011 п. н. В отличие от прокариот в эукариотических клетках одновременно активно транскрибируется от 1 до 10% ДНК. Состав транскрибируемых последовательностей и их количество зависят от типа клетки и стадии онтогенеза. Значительная часть нуклеотидных последовательностей у эукариот не транскрибируется вообще — молчащая ДНК.
Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей. Так, около 10% генома мыши составляют тандемно расположенные (друг за другом) короткие нуклеотидные последовательности, повторенные до 106 раз. Эти высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются. Около 20% генома мыши образовано умеренными повторами, встречающимися с частотой 103—105 раз. Такие повторы распределены по всему геному и транскрибируются в РНК. К ним относятся гены, контролирующие синтез гистонов, тРНК, рРНК и некоторые другие. Остальные 70% генома мыши представлены уникальными нуклеотидными последовательностями. У растений и амфибий на долю умеренно и высоко повторяющихся последовательностей приходится до 60% генома.
Избыточность генома эукариот объясняется также экзон-интронной организацией большинства эукариотических генов, при которой значительная часть транскрибированной РНК удаляется в ходе следующего за синтезом процессинга и не используется для кодирования аминокислотных последовательностей белков (см.
разд. 3.4.3.2).
Внастоящее время окончательно не выяснены функции молчащей ДНК, которая составляет значительную часть генома, реплицируется, но не транскрибируется. Высказывают предположения об определенном значении такой ДНК в обеспечении структурной организации хроматина (см. разд. 3.5.2.2). Некоторая часть нетранскрибируемых нуклеотидных последовательностей, очевидно, участвует в регуляции экспрессии генов (см. разд. 3.6.6).
Характеризуя наследственный материал прокариотической клетки в целом, необходимо отметить, что он заключен не только в нуклеоиде, но также присутствует в цитоплазме в виде небольших кольцевых фрагментов ДНК —
плазмид.
Плазмиды — это широко распространенные в живых клетках внехромосомные генетические элементы, способные существовать и размножаться в клетке автономно от геномной ДНК. Описаны плазмиды, которые реплицируются не автономно, а только в составе геномной ДНК, в которую они включаются в определенных участках. В этом случае их называют эписомами.
Впрокариотических (бактериальных) клетках обнаружены плазмиды, которые несут наследственный материал, определяющий такие свойства, как способность бактерий к конъюгации, а также их устойчивость к некоторым лекарственным веществам.
164
Вэукариотических клетках внехромосомная ДНК представлена генетическим аппаратом органелл — митохондрий и пластид, а также нуклеотидными последовательностями, не являющимися жизненно необходимыми для клетки (вирусоподобными частицами). Наследственный материал органелл находится в их матриксе в виде нескольких копий кольцевых молекул ДНК, не связанных с гистонами. В митохондриях, например содержится от 2 до 10 копий мтДНК.
Внехромосомная ДНК составляет лишь небольшую часть наследственного материала эукариотической клетки. Например, мтДНК человека содержит 16569 п.н.
ина её долю приходится менее 1% всей клеточной ДНК.
Вотличие от хромосомной ДНК, мтДНК характеризуется высокой «плотностью генов». В них нет интронов, а межгенные промежутки невелики. В кольцевой мтДНК человека содержится 13 генов, кодирующих белки (3 субъединицы цитохром С-оксидазы, 6 компонентов АТФазы и др.) и 22 гена тРНК. Значительная часть белков митохондрий и пластид синтезируется в цитоплазме под контролем геномной ДНК.
Если большинство ядерных генов представлены в клетках организма в двойной дозе (аллельные гены), то митохондриальные гены представлены многими тысячами копий па клетку.
Для генома митохондрий характерны межиндивидуальные различия, но в клетках одного индивида, как правило, мтДНК идентична.
Совокупность генов, расположенных в цитоплазматических молекулах ДНК, называют плазмоном. Он определяет особый тип наследования признаков — цитоплазматическое наследование (см. разд. 6.3.2.).
3.6.4.Эволюция генома
3.6.4.1.Геном предполагаемого общего предка про- и эукариот
Общие принципы организации наследственного материала, представленного нуклеиновыми кислотами, а также принципы записи генетической информации у про- и эукариот свидетельствуют в пользу единства их происхождения от общего предка, у которого уже была решена проблема самовоспроизведения и записи информации на основе репликации ДНК и универсальности генетического кода. Однако геном такого предка сохранял большие эволюционные возможности, связанные с развитием надмолекулярной организации наследственного материала, разных путей реализации наследственной информации и регуляции этих процессов.
Многочисленные указания на различия в организации генома, деталях процессов экспрессии генов и механизмов ее регуляции у про- и эукариот (см. §3.4; 3.5; 3.6) свидетельствуют в пользу эволюции названных типов клеток по разным направлениям после их дивергенции от общего предка.
Существует предположение, что в процессе возникновения жизни на Земле первым шагом явилось образование самовоспроизводящихся молекул нуклеиновых кислот, не несущих первоначально функции кодирования аминокислот в белках.
165
Благодаря способности к самовоспроизведению эти молекулы сохранялись во времени. Таким образом, первоначальный отбор шел на способность к самосохранению через самовоспроизведение. В соответствии с рассмотренным предположением позднее некоторые участки ДНК приобрели функцию кодирования, т.е. стали структурными генами, совокупность которых на определенном этапе эволюции составила первичный генотип. Экспрессия возникших кодирующих последовательностей ДНК привела к формированию первичного фенотипа, который оценивался естественным отбором на способность выживать в конкретной среде.
Важным моментом в рассматриваемой гипотезе является предположение о том, что существенным компонентом первых клеточных геномов была избыточная ДНК, способная реплицироваться, но не несущая функциональной нагрузки в отношении формирования фенотипа. Предполагают, что разные направления эволюции геномов про- и эукариот связаны с различной судьбой этой избыточной ДНК предкового генома, который должен был характеризоваться достаточно большим объемом. Вероятно, на ранних стадиях эволюции простейших клеточных форм у них еще не были в совершенстве отработаны главные механизмы потока информации (репликация, транскрипция, трансляция). Избыточность ДНК в этих условиях создавала возможность расширения объема кодирующих нуклеотидных последовательностей за счет некодирующих, обеспечивая возникновение многих вариантов решения проблемы формирования жизнеспособного фенотипа.
3.6.4.2. Эволюция прокариотического генома
По мере совершенствования и повышения надежности главных механизмов потока информации значение избыточной ДНК в повышении выживаемости организмов снижалась. В такой ситуации одним из возможных направлений изменения генома было уменьшение его размеров за счет утраты некодирующих нуклеотидных последовательностей. Именно так можно представить эволюционный путь, пройденный геномом современных прокариот. Одновременно в качестве механизмов, поддерживающих выживаемость этих форм, в историческом развитии закреплялось свойственное им короткое время генерации, т.е. интенсивное размножение и быстрая смена поколений (кишечная палочка делится каждые 20 мин). Перечисленные особенности хорошо сочетаются с гаплоидностью прокариот, что приводит к воспроизведению в фенотипе любой мутации.
Экспрессия 95% ДНК, относительно малые размеры генома, гаплоидность, проявление в фенотипе практически каждой мутации в сочетании с коротким временем генерации обусловливают высокую приспособленность. Вместе с тем для прокариотического типа организации не свойственны обширные и разнообразные изменения структуры. Вследствие этого описанное направление эволюции, обеспечивая высокую способность к выживанию (прокариоты существуют на Земле около 3,5 млрд. лет), является тупиковым в плане прогрессивной эволюции живых существ.
166
3.6.4.3.Эволюция эукариотического генома
Вотличие от изменений прокариотического генома преобразования генома в эволюции эукариот связаны с нарастающим увеличением количества ДНК. Это увеличение наблюдается в процессе прогрессивной эволюции эукариот (см. рис. 1.2
иразд. 3.6.3). На фоне такого увеличения большая часть ДНК является «молчащей», т.е. не кодирует аминокислот в белках или последовательностей нуклеотидов в рРНК и тРНК. Даже в пределах одного гена молчащие (интроны) и кодирующие (экзоны) участки могут перемежаться. В составе ДНК обнаруживаются высоко и умеренно повторяющиеся последовательности. Вся масса ДНК распределена между определенным числом специализированных структур —хромосом. Хромосомы в отличие от нуклеоида прокариот имеют сложную химическую организацию. Эукариоты в большинстве случаев диплоидны. Время генерации у них значительно больше, чем у прокариот. Отмечаемые особенности, оформившиеся в ходе эволюции генома эукариот, допускают широкие структурные изменения и обеспечивают не только адаптивную (приспособительную), но и прогрессивную эволюцию.
Среди перечисленных выше моментов увеличение размеров генома в эволюции эукариот привлекает особое внимание. Этот процесс может осуществляться различными способами. Наиболее резко размер генома изменяется в результате полиплоидизации, которая достаточно широко распространена в природе. Она заключается в увеличении количества ДНК и хромосом, кратном гапловдному. Достигаемое в результате состояние полиплоидии приводит к увеличению дозы всех генов и создает избыток «сырого» генетического материала, который впоследствии видоизменяется в результате мутаций и отбора.
По-видимому, в ходе эволюции в результате накопления мутаций и дивергенции нуклеотидных последовательностей полиплоидизация сопровождалась переходом к диплоидному состоянию. Само по себе увеличение дозы генов еще не означает достижения однозначно положительного биологического результата. Об этом свидетельствует развитие в эволюции эукариот механизмов компенсации возрастающей дозы генов в ходе их экспрессии путем сокращения времени жизни в клетках зрелой РНК. Так, у тетраплоидных карповых рыб в ответ на увеличение дозы генов рРНК в молекулах рРНК соматических клеток возникают скрытые внутренние разрывы, которые приводят к преждевременному их старению и сокращению содержания в цитоплазме.
Если бы увеличение объема генома происходило только в результате полиплоидизации, то в природе должно было бы наблюдаться скачкообразное изменение его размеров. На самом деле этот процесс демонстрирует плавное увеличение содержания ДНК в геноме. Это позволяет допустить возможность других механизмов, изменяющих его объем.
Действительно, некоторое значение в определении размера генома имеют хромосомные перестройки, сопровождающиеся изменением содержания ДНК в них,
167
такие, как дупликации, делении и транслокации. Они обусловливают повторение, утрату некоторых последовательностей в составе хромосомы или перенос их в другие хромосомы.
Важным механизмом увеличения объема генома является амплификация нуклеотидных последовательностей, которая заключается в образовании их копий, что приводит к возникновению повторяющихся участков ДНК. Особенностью генома эукариот является наличие таких повторов в большом количестве, свидетельствующее о существенном вкладе механизма амплификации в увеличение размеров наследственного материала. Амплифицированные последовательности образуют семейства, в которых они собраны вместе (тандемная организация) или же распределяются по разным хромосомам. Конкретные изменения, приводящие,, к амплификации, бывают различными. Появление тандемов повторяющихся последовательностей объясняется, например, неравным кроссинговером, вследствие которого возникают многократные дупликации отдельных участков ДНК. Возможна амплификация путем вырезания фрагмента с последующей его репликацией вне хромосомы и встраиванием копий в другие хромосомы. Предполагают также амплификацию, осуществляемую путем «обратной транскрипции» ДНК на РНК с участием фермента обратной транскриптазы с последующим встраиванием копий ДНК в различные локусы хромосом.
Во всех случаях амплификация некоторой последовательности приводит к возникновению в геноме более или менее многочисленных повторов и способствует некратному увеличению его объема. Наличие таких повторов в сочетании с мутационным процессом является предпосылкой дивергентной эволюции однотипных последовательностей в пределах семейства с соответствующим изменением свойств кодируемых белков или РНК.
Ярким примером эволюционной судьбы амплифицированных нуклеотидных последовательностей являются семейства глобиновых генов, широко распространенных в природе у видов разных уровней организации. У высших позвоночных известен ряд глобиновых генов, контролирующих синтез полипептидов гемоглобина. У человека в геноме имеется восемь активных глобиновых генов, образующих два семейства (см. рис. 6.5). Семейство генов, определяющих синтез α-глобинов, содержит ξ1-глобиновый ген, активно функционирующий в эмбриогенезе, и два α-глобиновых гена, которые экспрессируются у плода и взрослого человека. Это семейство генов располагается в 16-й хромосоме в следующем порядке: 5'—ξ2—ψξ1—α2—α1—3'. Семейство генов, определяющих синтез β-глобинов, расположенное в 11-й хромосоме, содержит ε- глобиновый ген эмбриона, два сходных γ-глобиновых гена плода GγAγ малый δ- и большой β-глобиновые гены взрослых: 5' — ε —Gγ — Aγ — δ — β — 3'.
Изучение гомологии продуктов указанных генов и генов миоглобина у разных видов организмов позволило предположить общность происхождения этих семейств. Вероятно, около 1100 млн. лет назад произошла дупликация генапредшественника, давшая начало гемоглобиновым и миоглобиновым генам. Позднее, около 500 млн. лет назад, на ранней стадии эволюции позвоночных
168
произошла дупликация, давшая начало двум (α и β) семействам глобиновых генов, сопровождавшаяся транслокацией. Примерно 200 млн. лет назад очередная дупликация привела к возникновению в семействе β-глобиновых генов генов β- глобинов плодов и взрослых. Около 100 млн. лет назад произошло образование ε- и γ-глобиновых генов и, наконец, 40 млн. лет назад появились δ- и β-глобиновые гены.
Семейства α- и β-глобиновых генов организованы в генные кластеры, возникшие, вероятно, в результате тандемной дупликации генов. В составе указанных кластеров наряду с активно функционирующими на разных стадиях онтогенеза генами обнаружены неактивные, или псевдогены. Последние возникли, вероятно, в результате появления в них изменений, несовместимых с врзможностью их экспрессии. В семействе β-глобиновых генов содержится два псевдогена: ψβ1 и ψβ2. В α-семействе имеются ψξ1 и ψξ2 псевдогены.
Дивергенция амплифицированных последовательностей с образованием разных генов или их семейств обусловлена накоплением в них различных изменений в виде замен оснований или других генных мутаций. О гомологии глобиновых генов обоих семейств свидетельствует наличие во всех активных глобиновых генах позвоночных двух интронных участков, занимающих в них строго одинаковое положение. Такую же организацию имеют и псевдогены ψα1 человека, ψα2 у кролика. Однако в ψα3-псевдогене мыши в ходе эволюции оба интрона оказались точно вырезанными.
Результатом амплификации небольших последовательностей ДНК в пределах функциональной единицы является удлинение гена, при котором из простых генов могут возникать более сложные. Это может происходить за счет тандемных дупликации. Например, в генах, кодирующих вариабельные участки иммуноглобулинов мыши, последовательности из 600 п.н. образуются в результате 12 тандемных повторов исходной предковой последовательности в 48 п.н. Другим примером удлинения гена посредством тандемных дупликации служит ген коллагена α2, который у курицы состоит из 34 000 п.н. и содержит больше 50 экзонов. Длина таких участков во всех случаях кратна девяти нуклеотидным парам. Эволюция этих экзонов, очевидно, шла от гипотетического исходного строительного блока длиной в 54 пары нуклеотидов.
Таким образом, амплификация нуклеотидных последовательностей, происходившая в процессе эволюции генома, обеспечивала не только его количественное увеличение, появление семейств генов, но и создавала предпосылки для накопления в них изменений, дивергенции генов, увеличения разнообразия контролируемых ими продуктов.
3.6.4.4. Подвижные генетические элементы
Определенная роль в эволюции геномов как про-, так и эукариотических клеток принадлежит так называемым подвижным генетическим элементам — транспозонам. Они представляют собой автономные единицы, несущие в нуклеотидной последовательности информацию о структуре особых белков,
169
которые обеспечивают их способность к перемещению из одного участка генома в другой. Такое перемещение — транспозиция — может происходить в строго определенные участки хромосом, узнаваемые этими специфическими белками. Транспозиция предполагает репликацию нуклеотидной последовательности подвижного генетического элемента и встраивание копии в ДНК-мишень с сохранением другой копии в прежнем месте.
Установлена также способность подвижных генетических элементов к точному вырезанию и удалению их из хромосомы. Перемещение таких нуклеотидных последовательностей в пределах генома может влиять на регуляцию экспрессии генов, которые прилежат к месту встраивания этих элементов. В результате таких перемещений могут активироваться ранее не активные гены, и наоборот.
Обнаружение подвижных генетических элементов в геномах как про-, так и эукариот указывает на определенные эволюционные преимущества, связанные с их наличием в наследственном материале. Возможно, рекомбинационные процессы, обеспечиваемые подвижными генетическими элементами, имеют немаловажное значение в структурной эволюции генома.
3.6.4.5.Роль горизонтального переноса генетического материала
вэволюции генома
Наряду с транспозонами, не способными очевидно, существовать вне генома и образовывать свободные молекулы ДНК, описаны элементы, обнаруживаемые как в составе генома, так и вне его. Существование таких подвижных элементов дает возможность обсуждать роль горизонтального переноса генетического материала в
эволюции генома.
Если описанные выше изменения структуры генома передаются из поколения в поколение организмов одного и того же вида, т.е. по вертикали, то горизонтальный перенос генетической информации может происходить и между организмами разных видов, одновременно существующими на Земле. В настоящее время доказана возможность изменения наследственных свойств у бактерий путем введения в бактериальную клетку чужеродной ДНК при конъюгации или с помощью фагов. Оказывается, чужеродную ДНК можно ввести и в эукариотическую клетку, где она будет сохраняться как внехромосомный элемент или интегрироваться в геном и экспрессироваться.
Недавно получены данные, свидетельствующие о том, что гены могут переходить от одного эукариотического организма к другому и даже от эукариот к прокариотам, хотя это происходит крайне редко. Примером могут служить данные о несовпадении скоростей эволюции отдельных последовательностей генов гистонов у некоторых видов морских ежей. Это можно объяснить относительно поздним по сравнению с временем дивергенции этих видов горизонтальным переносом указанных последовательностей, проявляющих большее сходство, чем этого можно было ожидать.
170