

Было установлено, что к поверхности ретикулоцита могут присоединяться25000—50000 молекул трансферрина, нагруженных железом. Меченый трансферрин легко присоединяется к ретикулоцитам, но не присоединяется к лейкоцитам, тромбоцитам и зрелым эритроцитам.
В плазме имеется небольшое количество белков, которые, помимо трансферрина, присоединяют железо. Однако структура этих белков и значение этого явления пока неясны.
Железо эритрокариоцитов
После того как трансферрин«разгружает» железо на поверхности эритрокариоцита, оно проникает внутрь клетки. Трансферрин в большинстве случаев возвращается в плазму, но некоторые молекулы трансферрина при этом проникают внутрь эритрокариоцита и связываются с молекулой носителя белком с молекулярной массой 20000. Bales, Workman (1974) описали белок сидерофилин, который связывает железо в клетке и передает его для синтеза гемоглобина и ферритина.
Железо, возможно в комплексе с трансферрином или сидерофилином, проникает в митохондрии, где происходит синтез гема из протопорфирина и железа. Судьба проникшего в клетку трансферрина неизвестна. Образование ферритина происходит в эритрокариоците из апоферритина, синтезируемого в клетке, и железа, проникшего в клетку. Предполагалось, что ферритин используется в эритрокариоците как обязательный или необязательный субстрат, участвующий в синтезе гемоглобина. Однако в настоящее время установлено, что синтез гемоглобина не требует обязательного включения ферритина, и нет пока убедительных данных о том, что ферритин в эритрокариоците может быть использован для синтеза гемоглобина. Наиболее вероятно, что синтез ферритина в эритрокариоците нужен для удаления из клетки избыточного железа, не вошедшего в гемоглобин. Этот ферритин собирается в лизо-сомах, а затем удаляется из клетки как в костном мозге, так и после удаления из клетки ядра в циркуляции. В процессе удаления гранул железа из циркулирующей клетки, повидимому, принимает участие селезенка, так как в эритроцитах людей с удаленной по поводу травмы селезенкой (без заболеваний системы крови) обнаруживаются гранулы железа, тогда как в норме их выявить в зрелых эритроцитах не удается.
Депонирование железа
Основным белком, используемым для сохранения избытка железа в организме, является ферритин — водорастворимый комплекс гидроокиси трехвалентного железа и белка апоферритина. Гидроокись железа соединена с остатком фосфорной кислоты.
Ферритин напоминает по форме грецкий орех, скорлупа ореха — это белок апоферритин, а внутри находятся в различном количестве атомы железа, почти вплотную прилегающие один к другому. Ферритин может вместить до 4500 атомов железа, практически одна молекула содержит около 3000 атомов. Молекулярная масса ферритина зависит от числа атомов железа, а этот показатель может колебаться. В среднем молекулярная масса ферритина близка к 460000. Ферритин в норме имеется в плазме и практически почти во всех клетках организма, но основные ткани, в которых он содержится, — это печень и мышцы.
Гемосидерин — белок, содержащий железо, обнаруживается в фагоцитирующих макрофагах и их производных, макрофагах костного мозга, макрофагах селезенки, купферовских клетках печени. Гемосидерин — это частично денатурированный и частично депротеинизированный ферритин. В гемосидерине нет белка, отличного от ферритина. Гемосидерин иммунологически полностью идентичен ферритину [Wohler, 1964]. Молекула ферритина содержит 20% железа, тогда как в гемосидерине содержание железа более высокое —25—30%. В отличие от ферритина гемосидерин нерастворим в воде.
Как гемосидерин, так и ферритин используется в качестве белков запаса, однако скорость использования гемосидерина значительно более медленная, чем ферритина. Hershko изучал вопросы регуляции обмена железа, которое находится в запасах. На схеме (рис. 1) показано, что железо запасов может быть как в паренхиматозных клетках, так и в фагоцитирующих макрофагах. В норме основную часть железа, связанного с трансферрином, организм использует для эритропоэза. Фагоцитирующие макрофаги, получившие железо при разрушении в них эритроцитов, в основном передают это железо трансферрину, который использует это железо вновь для эритропоэза. Паренхиматозные клетки тоже содержат железо, но это железо в основном в них откладывается, и лишь малая часть его передается трансферрину и используется для эритропоэза.
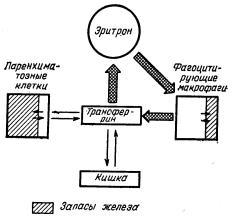
Рис. I. Схема внутреннего транспорта железа(по Hershko). В норме основная часть железа из трансферрина утилизируется эритрокариоцитами. Железо из нежизнеспособных эритроцитов утилизируется фагоцитирующими макрофагами. Паренхиматозные клетки получают железо от трансферрина. Основная часть железа, входящего в макрофаги, быстро обменивается, лишь малая часть остается в запасах; железо, входящее в паренхиматозные клетки, обменивается лишь незначительно.
Всвою очередь паренхиматозные клетки получают железо от трансферрина.
Вопытах на крысах установлено [Cook et al., 1972; Hershko et al., 1973], что в паренхиматозные клетки
проникает железо из сыворотки, связанное с трансферрином и входящее в состав ферритина сыворотки, а также содержащееся в гемоглобине (связанном с гаптоглобином сыворотки) или геме (связанном с гемопексином сыворотки). В то же время железо из эритроцитов, подвергнутых нагреванию для ускорения их гибели или из комплекса ферритина с антителами к ферритину, а также из введенного животным коллоидного железа поступает в основном в фагоцитирующие макрофаги. Макрофаги получают главным образом железо от нежизнеспособных эритроцитов. В макрофагах определяется выраженная активность фермента гемоксигеназы, который разрушает гемоглобин с образованием билирубина, окиси углерода и железа, способного присоединиться к трансферрину. В освобождении железа из макрофагов участвуют аскорбиновая кислота и фермент ферроксидаза. В 1966 г. Osaki с соавт. показали, что белок сыворотки, содержащий медь, — церулоплазмин — обладает ферроксидазной активностью, способен окислять железо для его соединения с трансферрином.
Паренхиматозные клетки, содержащие железо, располагаются главным образом в печени. В них хранится около 1/3 запасов железа. У здоровых людей за сутки в паренхиматозные клетки поступает0,09 мг железа на 100 мл крови. Это составляет около 1/5 части расхода трансферринового железа. При выраженном дефиците железа в организме поступление железа в паренхиматозные клетки печени уменьшается до0,01 мг на 100 мл крови в сутки, а при значительном избытке железа, апластических анемиях, почечной недостаточности — увеличивается до 0,45 мг на 100 мл крови в сутки [Hershko, 1977].
В отличие от железа макрофагов железо, находящееся в паренхиматозных клетках, расходуется медленно. В опытах на крысах было установлено, что после введения меченого ферритина из гепатоцитов расходуется за сутки 4,3% железа, тогда как из макрофагов — до 70% [Hershko et aL, 1974]. Cook с соавт. (1970) установили, что у здоровых людей за сутки из паренхиматозных клеток тратится 0,09 мг железа, а из макрофагов во много раз больше —0,41 мг. Аскорбиновая кислота увеличивает освобождение железа из макрофагов, но не влияет на освобождение его из гепатоцитов. При сочетании дефицита аскорбиновой кислоты с избыточным количеством железа в фагоцитирующих макрофагах содержание железа сыворотки низкое. Прием аскорбиновой кислоты приводит к подъему уровня железа сыворотки [Bothwell et al„ 1964].
Освобождение железа из паренхиматозных клеток увеличивается при кровотечениях и уменьшается при массивных гемотрансфузиях. При кровотечениях уменьшается захват эритроцитов макрофагами; следовательно, освобождение железа макрофагами в такой ситуации имеет меньшее значение.
Потери железа из организма
Раньше потери железа из организма пытались оценить химическими методами, однако они не годятся для этой цели, так как с калом теряется не только железо, проникшее туда с желчью или через эпителий кишки, но и железо пищи, которое не всосалось. Поэтому для оценки потери железа из организма используются радиологические методы исследования,
Finch (1959), исследовавший потерю железа с калом у мужчин в течение длительного периода, пришел к заключению, что мужчины теряют за сутки не менее 0,6 мг железа. Green с соавт. (1968) приводят более высокие цифры потери железа у мужчин: около 0,4 мг железа теряется с калом в виде крови, 0,25 мг железа выде-