

Поток информации в клетке.
Кодирование и реализация генетической информации.

Значение потока информации. Доказательства генетической роли ДНК.
Благодаря потоку информации клетки сохраняют и передают потомкам многовековой опыт своих предшественниц, а именно сохраняют индивидуальную и видовую специфичность, воссоздают свою структурную организацию и сохраняют способность к выполнению своих функций в организме.

ДНК была открыта в 1869 году швейцарским биохимиком Фридрихом Мишером в ядрах лейкоцитов (клетках гноя), но изучать ее строение начали в 20-е годы ХХ столетия.
В1924 году немецкий биохимик Хельген обнаружил, что в состав хромосом кроме гистоновых белков (55%) входит ДНК (до 40%).
В1933 году Коссель и Левин разложили молекулу ДНК и установили, что в ее состав входит 3 компонента: азотистое основание, углевод дезоксирибоза и остаток фосфорной кислоты.

Но наличие ДНК – это не доказательство ее генетической роли.
Первые доказательства роли ДНК в передаче наследственной информации были получены в 1928 году английским бактериологом Ф. Гриффитсом, изучившим явление трансформации. Трансформация – это способность одного штамма бактерий встраивать участки молекулы ДНК другого
штамма и приобретать при этом свойства последнего.

Строение, свойства и функции ДНК.
Биополимер, мономер – нуклеотид. 2 полинуклеотидных цепи, которые закручены вокруг общей оси. Нуклеотид состоит из 3 компонентов. Гетероциклические основания- это производные пурина (аденин и гуанин) и пиримидина (цитозин и тимин). Связь межу нуклеотидами сахарофосфатная, а между азотистыми основаниями слабые водородные связи (между Г и Ц -3, между А и Т -2).

Пространственная структура молекулы ДНК
расшифрована в 1953 году Дж. Уотсоном, Ф. Криком и М. Уилкинсоном. Этому предшествовали эксперименты Э. Чаргаффа, получившие названия «правил Чаргаффа»:
молярная масса пуриновых оснований (А+Г) равна массе пиримидиновых оснований (Ц+Т).
молярное содержание А равно Т, а Г равно Ц.
Соотношение оснований более изменчиво у микроорганизмов и растений, чем у животных.
Уотсоном и Криком было определено, что молекула ДНК – две спирально закрученные антипараллельные полинуклеотидные цепи (напротив конца 3' одной цепи располагается 5' конец другой). Строгое соответствие нуклеотидов друг другу в парных цепочках ДНК (А-Т, Г-Ц) называется комплементарностью. Расстояние между плоскостями пар оснований вдоль оси молекулы постоянно и составляет 3,4 ангстрем. Один виток включает 10 пар оснований и имеет длину по оси 34 ангстрема.

Свойства ДНК.
Редупликация (образование 2-х дочерних идентичных цепей). В 1957 году М. Дельбрук и Дж. Стент предложили 3 схемы удвоения молекул ДНК.
А) консервативная схема – исходная двойная спираль ДНК остается неизменной и целостной в процессе синтеза и строит новую двухцепочечную молекулу.

Б) полуконсервативная схема – цепи двойной спирали молекулы ДНК расходятся, не разрываясь, и каждая из одиночных цепей ДНК служит матрицей для образования комплементарной цепи. При этом в каждой дочерней молекуле ДНК одна цепочка материнская, а другая – вновь синтезированная.
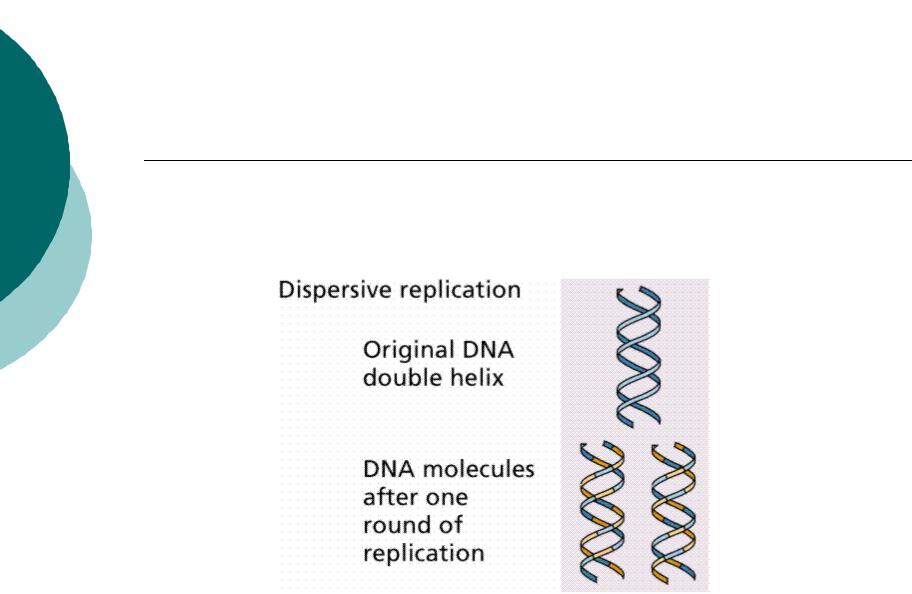
В) дисперсионная схема – в процессе удвоения молекулы ДНК составляющие ее цепи разрываются или разрушаются, так что после синтеза дочерних молекул последние включают в свой состав случайным образом перекомбинированные фрагменты исходящих молекул.
Наиболее аргументированной является полуконсервативная модель.

Редупликация осуществляется при участии ряда
ферментов. Хеликаза раскручивает и разделяет материнскую спираль ДНК на 2 нити, на которых по принципу комплементарности при участии фермента ДНК-полимеразы собираются дочерние цепи. Топоизомераза скручивает дочерние молекулы. Матричный синтез ДНК идет одновременно на обеих цепях материнской молекулы антипараллельно. Антипараллельность обеспечивает специфичность направления движения фермента ДНК-полимеразы, т.е. она может двигаться только в направлении 5'-3'. Цепочки собираются с разной скоростью. На лидирующей нити по мере раскручивания репликона постепенно и непрерывно наращивается дочерняя цепь. На отстающей нити дочерняя тепь синтезируется также в направлении 5'-3', но отдельными фрагментами (Оказаки) по мере раскручивания репликона. Фермент лигаза сшивает в единую нить фрагменты.