


Особую группу эфферентных нейронов спинного мозга составляют преганглионарные нейроны вегетативной нервной системы, расположенные как в боковых, так и в передних рогах серого вещества. Аксоны данных нейронов направляются к клеткам ганглиев симпатической цепочки и к интрамуральным ганглиям внутренних органов.
Вставочные, или промежуточные, нейроны спинного мозга представляют довольно разнородную группу нервных клеток, тела, дендриты, аксоны которых находятся в пределах спинного мозга. Вставочные нейроны имеют синаптические контакты только с другими нейронами и составляют значительное большинство нервных элементов спинного мозга.
Несмотря на то что различные вставочные нейроны значительно отличаются друг от друга размерами, характером дендритных разветвлений, аксональными проекциями и функциональными особенностями, им присущи и некоторые общие свойства. Как правило, вставочные нейроны имеют тело меньшего диаметра, чем ос-мотонейроны. Следовая гиперполяризация в большинстве разновидностей вставочных нейронов выражена незначительно. Поэтому они способны к генерации высокочастотной импульсной активности. Как видно на рис. 79, частота импульсации вставочного нейрона значительно превышает частоту импульсации мотонёйронов. Даже в ответ на одиночный стимул они обычно генерируют множественные разряды. Частота импульсов, регистрируемых в теле некоторых вставочных нейронов, может превышать 1000 в секунду.
К числу промежуточных нейронов относятся описанные выше клетки Реншоу (тормозные вставочные нейроны), получающие возбуждение от афферентных волокон мышечных рецепторов (проприоспинальные вставочные нейроны).
Тормозные вставочные нейроны включены в путь реципрокного торможения мотонейронов мышц-антагонистов.
Для проприоспинальных вставочных нейронов характерно свойство посылать длинные аксоны, проходящие через несколько сегментов и оканчивающиеся на мотонейронах. Вместе с тем на проприоспинальных клетках конвергируют волокна различных нисходящих трактов. Поэтому они являются важной релейной станцией на пути от вышележащих центров к мотонейронам. Рис. 80 демонстрирует пример моносинаптических ответов, возникающих в проприоспинальном нейроне при раздражении ретикулярной формации. Здесь же показана структура и местоположение этого нейрона.
Нейроны восходящих трактов также целиком находятся в пределах ЦНС. Тела этих клеток расположены в сером веществе спинного мозга, в то время как их аксоны проецируются к ядрам ростральных сегментов спинного мозга и к нейронам различных вышележащих образований. Наиболее хорошо изучены свойства нейронов дорсального и вентрального спинно-мозжечкового трактов, первые из которых, образуя так называемое грудное ядро (ядро Кларка), получают мощное возбуждающее влияние через афферентные пути мышечных рецепторов.
Наконец, центральные окончания первичных афферентных волокон характеризуются следующими особенностями. После вступления в спинной мозг афферентное волокно, как правило, дает начало восходящей и нисходящей ветвям, которые могут идти на значительные расстояния по спинному мозгу. Достигнув уровней конечного ветвления, колла-терали первичных афферентных волокон проникают в серое вещество и образуют концевые ветвления, которые заканчиваются синаптическими образованиями на телах и дендритах иннервируемых ими клеток. Концевые разветвления одного первичного афферентного волокна имеют многочисленные синапсы на одном мотонейроне. Кроме того, каждое афферентное волокно устанавливает синаптические контакты со многими мотонейронами. Установлено, что одно волокно, идущее от рецептора растяжения, образует синапсы, почти со всеми мотонейронами данной мышцы.
Терминали чувствительных афферентных волокон устанавливают синаптические контакты не только с дендритами и телами иннервируемых ими нейронов, но и сами получают сигнализацию от вставочных нейронов, включенных в цепь пресинаптического торможения, или от аксонов некоторых нисходящих систем.
Проводящие пути спинного мозга

Основные восходящие пути спинного мозга показаны на рис. 81 и в табл. 4. Часть из них представляет собой идущие без перерыва волокна первичных афферентных (чувствительных) нейронов. Эти волокна — тонкий (пучок Голля) и клиновидный (пучок Бурдаха) пучки идут в составе дорсальных канатиков белого вещества и заканчиваются в продолговатом мозге возле нейронных релейных ядер, называемых ядрами дорсального канатика, или ядрами Голля и Бурдаха. Волокна дорсального канатика являются проводниками кожно-механической чувст- рис. 81. Локализация основных восходящих путей в белом вительности. веществе спинного мозга (схема). Объяснение в тексте.

Остальные восходящие пути начинаются от нейронов, расположенных в сером веществе спинного мозга. Поскольку эти нейроны получают синаптические входы от первичных афферентных нейронов, их принято обозначать нейронами второго порядка, или вторичными афферентными нейронами. Основная масса волокон от вторичных афферентных нейронов проходит в составе латерального канатика белого вещества. Здесь расположен спиноталамический путь. Аксоны спиноталамических нейронов совершают перекрест и доходят не прерываясь через продолговатый и средний мозг до таламических ядер, где они образуют синапсы с нейронами таламуса. По спиноталамическим путям поступает импульсация от кожных рецепторов.
В латеральных канатиках проходят волокна спинно-мозжечковых трактов, дорсального и вентрального, проводящие в кору мозжечка импульсацию от кожных и мышечных рецепторов.
В составе латерального канатика идут и волокна спиноцервикального тракта, окончания которых образуют синапсы с релейными нейронами шейного отдела спинного мозга — нейронами
Рис.
82.
Локализация основных нисходящих путей
спинного мозга.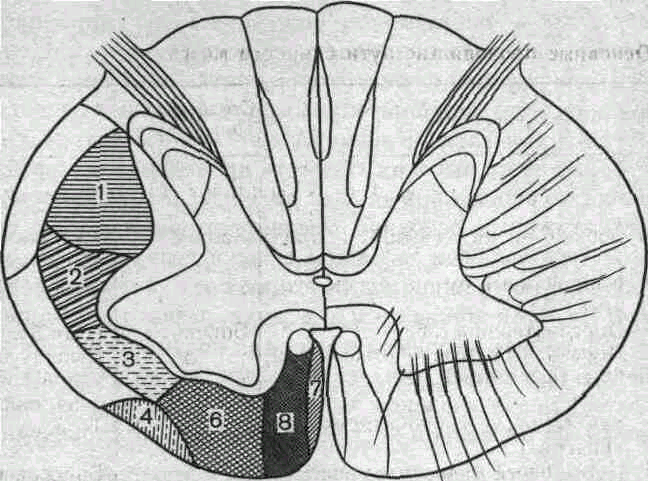
Путь болевой чувствительности локализуется в вентральных столбах белого вещества. Кроме того, в задних, боковых и передних столбах проходят собственные проводящие пути спинного мозга, обеспечивающие интеграцию функций и рефлекторную деятельность его центров.
Нисходящие пути спинного мозга также разделяются на несколько самостоятельных трактов, занимающих определенное положение в латеральных и вентральных канатиках белого вещества (рис. 82).
Эволюционно более древние нисходящие пути берут начало от нейронов, ядра которых расположены в пределах продолговатого мозга и моста. Это ретикулоспинальный и вестибулоспинальный тракты. Ретикулоспинальный тракт образован аксонами нейронов ретикулярной формации заднего мозга.
Ретикулоспинальные волокна идут в составе латеральных и вентральных канатиков спинного мозга и заканчиваются на многих нейронах серого вещества, в том числе на а- и y-мотонейронах. Сходную локализацию имеют волокна вестибулоспинального тракта, являющиеся главным образом аксонами нейронов латерального вестибулярного ядра, или ядра Дейтерса. Оба эти тракта не перекрещиваются.
Эволюционно более молодым нисходящим путем является руброспинальный тракт, достигающий наибольшего развития только у млекопитающих. Руброспинальные волокна являются аксонами нейронов красного ядра, расположенного в среднем мозге. Руброспинальный тракт совершает перекрест и идет в составе латеральных канатиков белого вещества.
Окончания руброспинальных волокон занимают в сером веществе спинного мозга более дорсальное положение, чем окончание волокон ретикуло- и вестибулоспинального трактов. Тем не менее часть из этих волокон образует-синапсы непосредственно на мотонейронах.
Наиболее важный нисходящий путь — кортико-спинальный, или пирамидный, тракт, нейроны которого расположены в двигательной зоне больших полушарий. Пирамидный тракт является Эволюционно самым молодым. Он появляется только у млекопитающих и наиболее развит у приматов и человека. Волокна пирамидного тракта совершают перекрест и идут в составе дорсолатеральных канатиков над руброспинальным трактом. Окончания кортико-спинальных волокон обнаруживаются главным образом на вставочных нейронах спинного мозга. Пирамидные аксоны, устанавливающие прямые связи с мотонейронами, относятся к миелинизированным волокнам большого диаметра и проводят импульсы с высокой скоростью.