

Миниатюрные постсинаптические потенциалы
Ацетилхолин секретируется двигательными нервными окончаниями не только при возбуждении, но и в покое. Различие состоит лишь в том, что в покое выделяются малые порции — «кванты» — ацетилхолина, а под влиянием нервного импульса в синаптическую щель одновременно выбрасывается значительное количество таких «квантов». «Квант» представляет собой «пакет» молекул медиатора в единичном пузырьке нервного окончания, изливающем свое содержимое в синаптическую щель. В концевой пластинке различных животных в каждом «кванте» содержится до 2000 молекул ацетилхолина. Выделение отдельных квантов в синаптическую щель в состоянии покоя вызывает кратковременную слабую деполяризацию постсинаптической мембраны мышечного волокна. Такая деполяризация получила название миниатюрного потенциала, поскольку она по своей амплитуде (0,5 мВ) в 50—80 раз меньше ПКП, вызываемого одиночным нервным импульсом. Миниатюрные потенциалы возникают обычно с частотой примерно один в секунду, они зарегистрированы не только в нервно-мышечных соединениях, но и в синапсах нервных клеток ЦНС.
Влияние кураре на нервно-мышечное соединение
Существует ряд веществ, также обладающих сродством к холинорецептору, но образующих с ним более прочную связь, чем ацетилхолин. К числу таких веществ относятся кураре и некоторые другие соединения (д-тубокурарин, диплацин, флакседил). После их воздействия на мышцу холинорецептор оказывается заблокированным и ни нервный импульс, ни искусственно введенный ацетилхолин не способны вызвать возбуждение мышечного волокна.
Многие годы изучение действия кураре на нервно-мышечную передачу представляло только теоретический интерес, и физиологи были очень далеки от мысли, что этот препарат может когда-либо найти применение в медицинской практике. Однако в связи с развитием хирургии возникла необходимость изыскания средств, которые позволили бы проводить оперативные вмешательства в условиях выключения естественного дыхания. И здесь кураре и его производные оказали большую помощь. В настоящее время многие полостные операции проводят в условиях искусственного дыхания на фоне нервно-мышечной блокады препаратами, действующими подобно кураре.
Исключительно прочную связь с холинорецептором образует токсин из яда змеи — (a-бунгаротоксин. Этот токсин, снабженный радиоактивной меткой, позволил выделить холинорецептор из мембраны. Химический анализ холинорецептора показал, что холинорецептор является липопротеидом с молекулярной массой около 300 000.
Холинэстераза и ее роль в процессах нервно-мышечной передачи
Установлено, что в области нервно-мышечного соединения в больших концентрациях присутствует фермент холинэстераза, способная быстро расщеплять ацетилхолин, выделяющийся в нервном окончании. Значение этого процесса становится ясным, если учесть, что в естественных условиях к мышце поступают быстро следующие друг за другом нервные импульсы и постсинаптическая мембрана, деполяризованная предшествующей порцией ацетилхолина, становится малочувствительной к действию следующей порции. Чтобы идущие друг за другом нервные импульсы могли осуществлять нормальное возбуждающее действие, необходимо к моменту прихода каждого из них «убрать» предшествующую порцию медиатора. Эту функцию и выполняет холинэстераза. Холин, освобождающийся при расщеплении молекул ацетилхолина, переносится обратно в нервное окончание специальной транспортной системой, существующей в пресинаптической мембране.
Существует ряд фармакологических агентов, обладающих способностью резко угнетать активность холинэстеразы. Их называют ингибиторами. К числу таких веществ
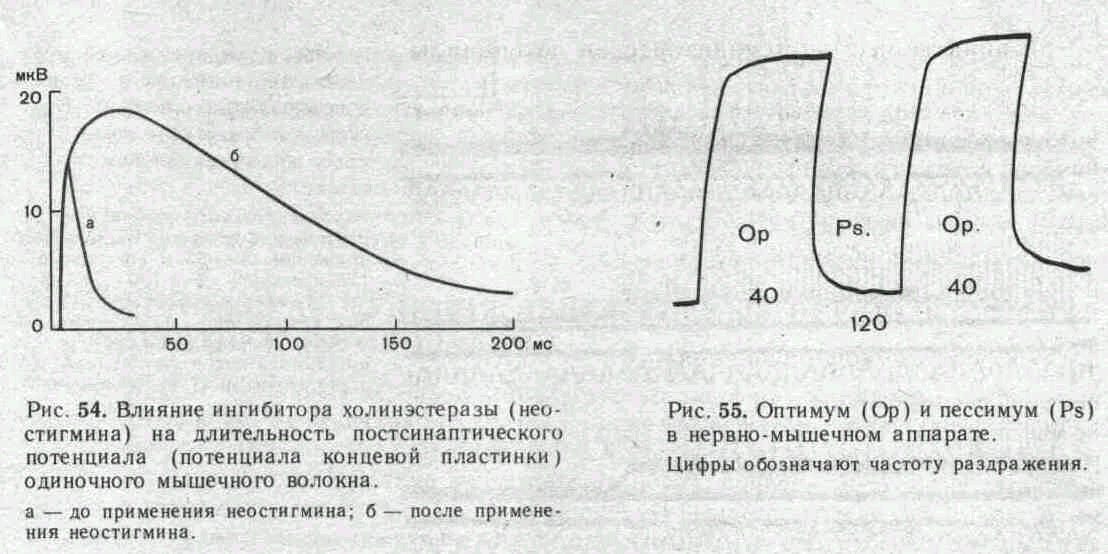
относятся эзерин, простигмин. галантамин. Если на нервно-мышечное соединение действует какое-либо из этих веществ, постсинаптический потенциал увеличивается по своей амплитуде и резко растягивается по времени.
Иллюстрацией этого является приведенная на рис. 54 запись ПКП, зарегистрированного в нервно-мышечном соединении лягушки до (а) и после (б) воздействия на мышцу вещества, угнетающего активность холинэстеразы.
При действии ингибитора холинэстеразы ритмическое раздражение нерва вызывает выраженную суммацию ПКП, что ведет к стойкой деполяризации постсинаптической мембраны и блоку проведения импульсов с нервного волокна на мышечное. При этом стойкая деполяризация постсинаптической мембраны приводит соседние участки мышечного волокна в состояние угнетения, обусловленное инактивацией натриевой и стойким повышением калиевой проводимости мембраны (состояние «католической депрессии»).
Следует отметить, что и в отсутствие ингибиторов холинэстеразы при условии очень частого раздражения нерва постсинаптические потенциалы (ПКП), вызываемые каждым нервным импульсом, суммируются, поскольку в межимпульсный интервал холинэстераза не успевает полностью расщепить выделяющийся в нервном окончании ацетилхолин. В результате суммации потенциалов постсинаптическая мембрана все более и более деполяризуется.