

2 курс / Гистология / Основы_гистологии_УЧЕБНО_МЕТОДИЧЕСКОЕ_ПОСОБИЕ_1
.pdfхроматофильная субстанция. После достижения растущим отростком нервной клетки рабочего органа вокруг новообразованного осевого цилиндра леммоциты образуют миелиновую оболочку.
Успешная регенерация нервных волокон зависит от многих факторов. Замедляют или полностью останавливают регенерацию: 1) наличие в зоне повреждения мертвых тканей, которые стимулируют разрастание здесь рубцовой ткани; 2) большое расстояние между отрезками нервного волокна; 3) сильное повреждение сосудов и нарушение кровоснабжения нерва. Все это стимулирует рубцевание. Разрастание рубцовой ткани иногда вызывает развитие ампутационной невромы, состоящей из разросшихся отростков нейронов и глии, окруженных грубой руб-цовой тканью. Невромы могут вызывать сильные (фантомные) боли и требуют оперативного вмешательства. Поэтому для улучшения срастания нервов производят тщательное сшивание (под операционным микроскопом) отрезков нерва, кровеносных сосудов, кровоснабжающих нерв, иссекают мертвые ткани. Стимулируется регенерация фактором роста нервов, который вырабатывается слюнными железами (зализывание ран животными имеет большой физиологический смысл), простатой. Стимулируют регенерацию нервных волокон анаболические гормоны, витамины, особенно витамин В12, фолиевая кислота, оротат калия, препараты ДНК и РНК и др.
В центральной нервной системе регенерация нервных волокон не наблюдается. Это связано не с отсутствием способности нейроцитов ЦНС к восстановлению отростков, а с образованием глиального рубца, нарушающего рост отростков нейроцитов.
Глава 14
НЕРВНЫЕ ОКОНЧАНИЯ. СИНАПСЫ. РЕФЛЕКТОРНЫЕ ДУГИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ НЕЙРОННОЙ ТЕОРИИ
Нервными окончаниями называются концевые разветвления отростков нервных клеток, в которых нервный импульс или генерируется, или передается на другую клетку (структуру). Все нервные окончания по функции делятся на три группы:
1.Эффекторные нервные окончания.
2.Рецепторные, или афферентные, нервные окончания.
3.Межнейронные синапсы.
ЭФФЕКТОРНЫЕ НЕРВНЫЕ ОКОНЧАНИЯ. Их функцией является вызывание эффекта, в зависимости от которого они делятся на две группы: 1) двигательные и 2) секреторные. Двигательные окончания подразделяются на: 1) двигательные окончания в скелетной мышечной ткани и 2)
двигательные нервные окончания в гладкой мышечной ткани.
https://t.me/medicina_free

Двигательные нервные окончания в скелетной мышечной ткани называются нервно-
мышечным синапсом, или моторной бляшкой (рис. 14.1,14.2). Представляют собой окончания аксонов мотонейронов передних рогов спинного мозга на поперечнополосатых мышечных волокнах. При образовании моторной бляшки миелиновое нервное волокно, подходя к мышечному волокну, теряет миелиновую оболочку. Осевой цилиндр внедряется в мышечное волокно, прогибая сарколемму. Терминальное ветвление аксона имеет на конце утолщение. Это так называемый нервный полюс нервно-мышечного синапса. В нем обнаруживаются митохондрии,
микротрубочки (нейротрубочки), синаптические пузырьки с нейромедиатором ацетилхолином.
Размеры последних составляют около 50 нм. В пресинаптической мембране есть утолщения — активные зоны, где происходит выделение(секреция) медиатора.
Плазмолемма мышечного волокна и прилегающая саркоплазма образуют мышечный полюс. Между пресинаптической и постсинаитической мембранами находится синаптическая щель. Она имеет ширину 50—100 нм и содержит базальную мембрану и отростки глиоцитов. На базалыюй мембране имеются сигнальные белки агрин, S-ламинин и др., которые служат метками, при
https://t.me/medicina_free

помощи которых регенерирующий аксон мотонейронов находит синапти-ческую зону на мышечном волокне.
Постсинаптическая мембрана имеет много складок, которые образуют вторичные синаптические щели. Они во много раз увеличивают поверхность синаптической щели и содержат материал базальной мембраны. В по-стсинаптической мембране имеются никотиновые холи-норецепторы, концентрация которых достигает 20—30 тыс. на 1 мкм2. В зоне синапса мышечное волокно теряет исчерченность (мио-фибриллы лежат глубже зоны синапса), но содержит большое количество митохондрий, профилей гранулярной ЭПС, рибосом, скопление ядер.
МЕХАНИЗМ РАБОТЫ НЕРВНО-МЫШЕЧНОГО СИНАПСА (см. рис. 12.10). Нервный импульс доходит до пресинантического полюса и вызывает увеличение мембранной проницаемости этого полюса для ионов кальция. При этом концентрация кальция в пресинаптическом полюсе резко возрастает благодаря: 1) высвооождению его из депо (гладкой ЭПС, митохондрий), а также 2) поступлению из внеклеточной среды. Далее кальций вызывает взаимодействие компонентов цитоске-лета, которые содержатся в пресинаптическом полюсе. Очевидно, наиболее выражены кинезиновый и динеиновый механизмы транспорта синаптичес-ких пузырьков к пресииаптической мембране.
https://t.me/medicina_free
После перемещения синаптических пузырьков к пресииаптической мембране происходит слияние с ней мембран, окружающих пузырьки, а затем пузырьки раскрываются в синаитическую щель и < выделяют в нее медиатор. Далее медиатор мигрирует к иостсинаптичес-кой мембране и вызывает ее деполяризацию, сливаясь с рецепторами ацетилхолина.
Деполяризация плазмолеммы мышечного волокна передается по Т-трубочкам на всю толщину
мышечного волокна, а затем с Т-тру-бочек переходит на терминальные |
цистерны |
сарконлазматического ретикулума (СПР). Это вызывает увеличение проницаемости |
СПР для |
ионов Са2+, который выходит из СПР и мигрирует к актиновым филаментам. Там он вызывает конформационные изменения в молекуле тропонина и открывает активные центры на ак-
активные центры >ш сатиновых филаментах. С этими центрами начинают связываться головки миозина. Происходит мышечное сокращение.
В иостсинаптической мембране содержится фермент ацетилхолшюсте-раза, который разрушает избыток ацетилхолина в синаптической щели и уменьшает время действия медиатора. Это необходимо для предотвращения перевозбуждения иостсинаптической мембраны.
Патология нервно-мышечного синапса. При отравлении фосфорорга-ническими соединениями (ФОС), которые относятся к боевым отравляющим веществам и широко используются в быту как инсектициды, актин ность ацетилхолинэстеразы подавляется. При этом в синаптической щели накапливается медиатор, вызывающий перевозбуждение иостсинаптичес-кой мембраны. В результате возникают судорожные сокращения мышц, затем сменяющиеся параличом. От паралича межреберных мышц больной погибает. Для лечения отравлений ФОС применяют реактиваторы ацетилхолинэстеразы, которые восстанавливают активность холинэстеразы и работу нервно-мышечного синапса.
Блокада ацетилхолиновых рецепторов на постсинаптической мембране может быть осуществлена некоторыми ядами (яд кураре). При этом становится невозможной передача возбуждения с нервного окончания на мышцу, и мышца полностью расслабляется. Синтетические аналоги кураре (курареподобные вещества, миорелаксанты) используются в хирургической практике для расслабления мышц при операциях. Существует заболевание миастения гравис, при котором в результате аутоиммунной реакции происходит разрушение ацетилхолиновых рецепторов в постсинаптической мембране нервно-мышечного синапса. Характеризуется прогрессирующей мышечной слабостью.
Двигательные нервные окончания на гладких мышцах представляют собой варикозные расширения терминалей аксона, которые контактируют с одним из миоцитов в миоцитарном комплексе. Терминали содержат си-наптические пузырьки с ацстилхолином или норадреналином.
Секреторные нервные окончания представляют собой терминали аксонов, которые вступают в тесную связь с секреторными клетками: или подходят к ним, не проникая через базальную мембрану, или пенетрируют ба-зальиую мембрану и вдавливаются в секреторные клетки, образуя терминальные расширения. Нейролемма аксона и плазмолемма секреторной клетки образуют соответственно пре- и постсинаптические мембраны, разделенные узкой синаптической щелью. Медиатор, выделившийся из синаптичес-ких пузырьков, вызывает деполяризацию мембраны секреторной клетки, что приводит к высвобождению кальция из депо (обычно он находится в митохондриях и секреторных гранулах). Кальций связывается с белком кальмо-дулином, и этот
https://t.me/medicina_free
комплекс вызывает два эффекта: полимеризацию микротрубочек и взаимодействие актиновых и миозиновых филамент, что способствует продвижению секреторных пузырьков к цитолемме, слиянию их мембраны с цитолеммой и ведет к последующему выделению секрета из клетки.
ЧУВСТВИТЕЛЬНЫЕ НЕРВНЫЕ ОКОНЧАНИЯ (РЕЦЕПТОРЫ)
Представляют собой терминальные разветвления дендритов нейроцита. КЛАССИФИКАЦИЯ. Существует несколько принципов классификации рецепторных нервных окончаний.
1.По месту восприятия раздражителя. Рецепторные нервные окончания делятся на три группы: экстерорецепторы, воспринимающие раздражение из внешней среды; интерорецепторы, служащие для восприятия раздражений из внутренней среды организма; проприорецепторы, воспринимающие информацию от опорно-двигательного аппарата.
2.В зависимости от специфичности раздражения, воспринимаемого рецептором. Выделяют:
механорецепторы, воспринимающие механические раздражители, перемещения частей тела; хеморецепторы воспринимают химические раздражители; терморецепторы улавливают изменения температуры, а ноцирецепторы воспринимают чувство боли.
3.По способу восприятия раздражителя выделяют контактные рецепторы, приходящие в состояние возбуждения при непосредственном воздействии на участок тела, и дистантные рецепторы, воспринимающие раздражитель, удаленный от организма (рецепторные клетки сетчатки глаза, органа слуха, обоняния).
4.Морфологическая классификация. В зависимости от строения все рецепторы делят на свободные и несвободные. Свободные рецепторные нервные окончания состоят только из конечных ветвлений дендрита чувствительного нейрона, а несвободные нервные окончания кроме терминалей нервного отростка имеют также и клетки олигодендроглии (леммоци-ты), которые окружают терминали дендрита и участвуют в восприятии раздражения. В свою очередь, несвободные нервные окончания делятся на неинкапсулированные (не окруженные по периферии соединительнотканной капсулой) и инкапсулированные, имеющие такую капсулу. Свободные нервные окончания воспринимают в основном болевые раздражения. Большинство несвободных нервных окончаний являются механорецепто-рами. В последнее время, однако, выдвигается небеспочвенная точка зрения, что не существует разделения рецепторов в зависимости от вида воспринимаемого раздражения, все рецепторы способны воспринимать раздражители любой модальности, а характер ощущения зависит от силы раздражителя.
МОРФОЛОГИЯ РЕЦЕПТОРОВ. 1. Свободные нервные окончания. Внаибольшем количестве представлены в коже. Это механорецепторы на волосяных фолликулах, ноцицептивные (воспринимающие болевые раздражители) нервные окончания в эпидермисе (рис. 14.3). Их много также в многослойном плоском неороговевающем эпителии, серозной оболочке. В эпидермисе они представлены древовидными ветвлениями дендритов псевдоуниполярных нейронов спинальпых ганглиев.
2. Несвободные неинкапсулированные нервные окончания представлены осязательными дисками Меркеля, а также нервными окончаниями соединительной ткани. Особенно много их в дерме. Осязательные диски Меркеля (рис. 14.3 б) состоят из отростка нервной клетки, который заканчивается расширением в виде П. диска. Этот диск образует синапс с клеткой Меркеля, которая лежит в эпидермисе. В цитоплазме клетки Меркеля есть секреторные гранулы с
https://t.me/medicina_free

нейромедиа тором. Механическое раздражение вызывает выделение гранул из клеток Меркеля, их содержимое ведет к деполяризации отростка нейроцита.
Несвободные неинкапсули-рованные окончания в соединительной ткани построены следующим образом. Осевой цилиндр освобождается от миелина и на значительном расстоянии окружается глиальными клетками, тесно с ними контактируя. Очень часто на поперечном разрезе видна билатеральная симметрия таких окончаний.
3. Несвободные инкапсулированные нервные окончания построены по общему принципу. К
этим окончаниям относятся нервные окончания в соединительной и мышечных тканях. Есть следующие разновидности этих окончаний: пластинчатые тельца Фатер-Пачини, осязательные
тельца Мейснера, концевые колбы Краузе, генитальные тельца Догеля, тельца Руффини, не- рвно-мышечные и нервно-сухожильные веретена и др.
https://t.me/medicina_free

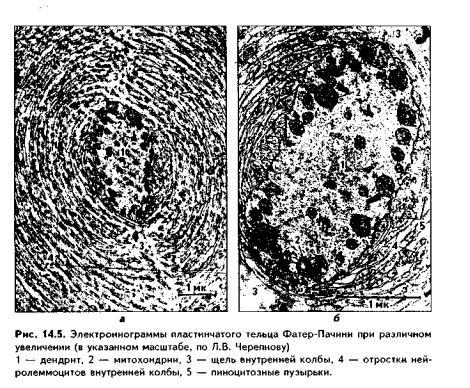
Наиболее распространены пластинчатые тельца Фатер-Пачини. Они встречаются в коже, молочной железе, в брыжейке, во внутреннил органах, около кровеносных сосудов, около суставов. Это крупные образования диаметром от 1 до 5 мм (рис. 14.4, 14.5). Имеют овальную форму и состоят из соединительнотканной капсулы, терминалей дендрита псевдоуниполярного нейрона и нейролеммоцитов (олигодендроглии). Дендрит при подходе ккапсуле теряет миелиновую оболочку и со всех сторон окружается нейролем-моцитами. Они формируют так называемую внутреннюю колбу. Эта колба снаружи покрыта соединительнотканной капсулой, которая часто называется наружной колбой. Капсула состой! из послойно параллельно лежащих кол-лагеновых волокон (образуют от 10 до 60 слоев) и клеток фиброцитов. В наружной капсуле встречаются кровеносные сосуды. Между наружной и внутренней колбами лежат специализированные отростчатые олигодендроглиоци-ты, контактирующие с осевым цилиндром. При давлении на тельце механическое воздействие во много раз усиливается слоями наружной колбы, что делает этот рецептор очень чувствительным. Давление смещает наружную колбу по отношению к внутренней. При этом раздражаются отростчатые олигоденд-роциты, передающие возбуждение на дендрит.
https://t.me/medicina_free
В сосочковом слое дермы обнаруживаются осязательные тельца Мейс-нера. Они являются механорецепторами и по размеру меньше телец Фа-тер-Пачини (50—140 мкм). Имеют овальную форму (рис. 14.3 и 14.6). Снаружи находится очень тонкая слоистая капсула — наружная колба. Дендрит псевдоуниполярного нейрона теряет миелиновую оболочку, разветвляется, и его ветви входят внутрь капсулы по спирали. Перпендикулярно к ним лежат глиальные клетки, которые вместе с терминалами дендритов образуют внутреннюю колбу. Незначительная деформация капсулы передается глиоцитам, которые имеют синаптическую связь с дендритом.
Концевые колбы Краузе являются барорецепторами и терморецепторами. Они лежат в дерме кожи, слизистых оболочках. Имеют небольшие (40—150 мкм) размеры. Также состоят из наружной капсулы и внутренней колбы. Внутренняя колба образована плоскими глиоцитами, между которыми проходят, формируя своеобразный клубочек, тонкие ветви дендрита. Наружная капсула очень тонкая.
Генитальные тельца Догеля находятся в особо чувствительных областях кожи, в первую очередь, в области наружных половых органов, коже молочных желез. Они похожи по строению на колбы Краузе, но в в отличие от них в тельце входят несколько отростков от нескольких нейро-цитов. Поэтому раздражение генитального тельца вызывает сильную иррадиацию возбуждения.
Тельца Руффини находятся в соединительной ткани кожи и в капсулах суставов. Воспринимают чувство давления. Имеют вид верстеновид-ных образований длиной до 2 мм. Осевой цилиндр во внутренней колбе разветвляется с образованием большого количества ветвей с булавовидными утолщениями на конце. Капсула хорошо выражена.
https://t.me/medicina_free

Вгладкой мышечной ткани чувствительные нервные окончания также инкапсулированы, они контактируют с группой гладких миоцитов.
Вскелетной мышечной ткани чувствительные нервные окончания называются нервно-
мышечными веретенами. Представляют собой инкапсулированные нервные окончания (рис. 14.7, 14.8). Наружная соединительнотканная капсула нервно-мышечного веретена окружает несколько тонких так называемых интрафузальных мышечных волокон. В отличие от обычных мышечных волокон, лежащих снаружи и называемых экстрафузаль-ными, интрафузальные волокна тонкие, содержат мало миофибрилл и имеют светлую цитоплазму. Различают два вида интрафузальных мышечных волокон (рис. 14.8). 1. ЯС-волокна. Ядра этих волокон лежат в центральной части мышечного волокна, образуя скопление в виде ядерной сумки (сокращенно ЯС). В месте расположения ядер волокно резко расширяется.
2. ЯЦ-волокна. Эти волокна имеют равномерную толщину, а ядра лежат но всей длине волокна в его центре, формируя ядерную цепь.
Вокруг данных двух видов интрафузальных волокон в их центральной части образуются специфические синапсы дендритов чувствительных нейронов в виде:
1) аннулоспиральных (кольцеспиральных) окончаний, в которых отростки нервных клеток закручены вокруг центральной части интрафузалыю-го волокна по спирали и на большом протяжении вступают в синаптичес-кую связь с ним; аннулосниральные окончания имеются как на ЯС-, так и на ЯЦ-волокнах. 2) гроздьевидных окончаний, которые находятся только на ЯЦволокнах. При этом они формируются не в центральной части, а на периферии волокна.
https://t.me/medicina_free

На интрафузальных волокнах имеются также двигательные нервные окончания, которые представлены аксонами у-мотонейронов передних рогов спинного мозга. Они регулируют длину интрафузальных волокон и поддерживают их тонус. Все свободное пространство между мышечными волокнами заполнено жидкостью и ограничено тонкой капсулой. Всякое изменение тонуса мышцы ведет к изменению давления жидкости в полости капсулы. При этом давление передается на дендриты. Аннулосниральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, а гроздьевидные — только на изменение длины. Благодаря нервно-мышечным веретенам организм по-
стоянно получает информацию о степени сокращения мышц, что формирует представление о положении тела в пространстве.
МЕЖНЕЙРОННЫЕ СИНАПСЫ
Это особый вид нервных окончаний, когда разветвления отростков одних нервных клеток заканчиваются на других нервных клетках. При помощи синапсов возбуждение передается с одной нервной клетки на другую.
Классификация синапсов. Существует несколько подходов к классификации синапсов.
https://t.me/medicina_free