

мембранных ферментов — цитохрома Р450, цитохром Р450 редуктазы, цитохром b5 редуктазы и цитохрома b5.
Аппарат Гольджи
Аппарат Гольджи — важная мембранная органелла, отвечающая за модификацию, накопление, сортировку и направление различных веществ в соответствующие внутриклеточные компартменты, а также за пределы клетки. Специфические ферменты мембраны комплекса Гольджи, гликозилтрансферазы, гликозилируя белки по остаткам серина, треонина или амидной группе аспарагина, завершают образование сложных белков — гликопротеинов.
Митохондриальные мембраны
Митохондрии — органеллы, окружённые двойной мембраной, специализирующиеся на синтезе АТФ путём окислительного фосфорилирования. Отличительная особенность внешней митохондриальной мембраны — содержание большого количества белка порина, образующего поры в мембране. Благодаря порину внешняя мембрана свободно проницаема для неорганических ионов, метаболитов и даже небольших молекул белков (меньше 10 кД). Для больших белков внешняя мембрана непроницаема, это позволяет митохондриям удерживать белки межмембранного пространства от утечки в цитозоль.
Для внутренней мембраны митохондрий характерно высокое содержание белков, около 70%, которые выполняют в основном каталитическую и транспортную функции. Транслоказы мембраны обеспечивают избирательный перенос веществ из межмембранного пространства в матрикс и в обратном направлении, ферменты участвуют в транспорте электронов (цепи переноса электронов) и синтезе АТФ.
Мембрана лизосом
Мембрана лизосом играет роль «щита» между активными ферментами (более 50), обеспечивающими реакции распада белков, углеводов, жиров, нуклеиновых кислот, и остальным клеточным содержимым. Мембрана содержит уникальные белки, например, АТФ-зависимую протонную помпу (насос), которая поддерживает кислую среду (pH 5), необходимую для действия гидролитических ферментов (протеаз, липаз), а также транспортные белки, позволяющие продуктам расщепления макромолекул покидать лизосому. Большинство белков лизосомальной мембраны сильно гликозилированы, углеводные составляющие, находящиеся на внутренней поверхности мембраны, защищают их от действия протеаз.
Строение и состав мембран
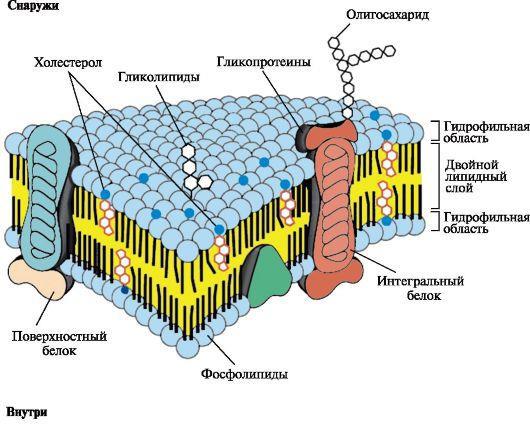
Биологические мембраны построены из липидов и белков, связанных друг с другом с помощью нековалентных взаимодействий. Основу мембраны составляет двойной липидный слой, в состав которого включены белковые молекулы (рис.1).
Липидный бислой образован двумя рядами амфифильных молекул, гидрофобные «хвосты» которых спрятаны внутрь, а гидрофильные группы - полярные «головки» обращены наружу и контактируют с водной средой.
Рис.1Поперечный разрез плазматической мембраны
Липиды мембран. В состав липидов мембран входят как насыщенные, так и ненасыщенные жирные кислоты. Ненасыщенные жирные кислоты встречаются в два раза чаще чем насыщенные, что определяет текучесть мембран и конформационную лабильность мембранных белков.
В мембранах присутствуют липиды трех главных типов - фосфолипиды, гликолипиды и холестерол (рис. 2,3,4). Чаще всего встречаются глицерофосфолипиды - производные фосфатидной кислоты.
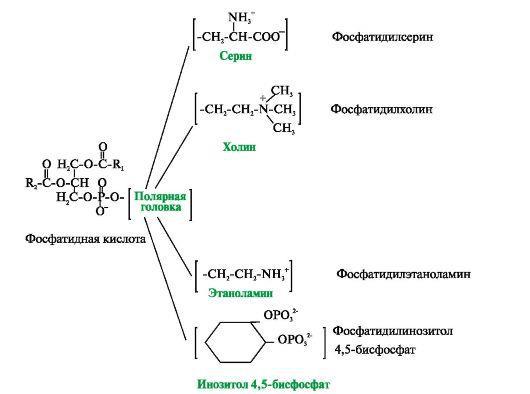
Рис.2 Глицерофосфолипиды.
Фосфатидная кислота - это диацилглицеролфосфат. R1, R2 - радикалы жирных кислот (гидрофобные «хвосты»). Со вторым углеродным атомом глицерола связан остаток полиненасыщенной жирной кислоты. Полярной «головкой» является остаток фосфорной кислоты и присоединенная к нему гидрофильная группа серина, холина, этаноламина или инозитола. Существуют также липиды -
производные аминоспирта сфингозина.
Аминоспирт сфингозин при ацилировании, т.е. присоединении жирной кислоты к NH2-группе, превращается в церамид. Церамиды различаются по остатку жирной кислоты. С ОН-группой церамида могут быть связаны разные полярные группы. В зависимости от строения полярной «головки» эти производные разделены на две группы - фосфолипиды и гликолипиды. Строение полярной группы сфингофосфолипидов (сфингомиелинов) сходно с глицерофосфолипидами. Много сфингомиелинов содержится в составе миелиновых оболочек нервных волокон. Гликолипиды представляют собой углеводные производные церамида. В зависимости от строения углеводной составляющей различают цереброзиды и ганглиозиды.
Холестерол содержится в мембранах всех животных клеток, он придает мембранам жесткость и снижает их жидкостность (текучесть). Молекула холестерола располагается в гидрофобной зоне мембраны параллельно

гидрофобным «хвостам» молекул фосфо- и гликолипидов. Гидроксильная группа холестерола, как и гидрофильные «головки» фосфо- и гликолипидов,
Рис.3 Производные аминоспирта сфингозина.
Церамид - ацилированный сфингозин (R1 - радикал жирной кислоты). К фосфолипидам относятся сфингомиелины, у которых полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Гидрофильной группой (полярной «головкой») гликолипидов является углеводный остаток. Цереброзиды содержат моно - или олигосахаридный остаток линейного строения. В состав ганглиозидов входит разветвленный олигосахарид, одним из мономерных звеньев которого является НАНК - N-ацетилнейраминовая кислота обращена к водной фазе. Молярное соотношение холестерола и других липидов в мембранах равно 0,3-0,9. Самое высокое значение имеет эта величина для цитоплазматической мембраны.
Увеличение содержания холестерола в мембранах уменьшает подвижность цепей жирных кислот, что влияет на конформационную лабильность мембранных белков и снижает возможность их латеральной диффузии. При повышении текучести мембран, вызванном действием на них липофильных веществ или перекисным окислением липидов, доля холестерола в мембранах возрастает.
Рис.4 Положение в мембране фосфолипидов и холестерола.
Молекула холестерола состоит из жесткого гидрофобного ядра и гибкой углеводородной цепи. Полярной «головкой» является ОН-группа у 3-го углеродного атома молекулы холестерола. Для сравнения на рисунке представлено схематическое изображение фосфолипида мембран. Полярная головка этих молекул значительно больше и имеет заряд
Липидный состав мембран различен, содержание того или другого липида, повидимому, определяется разнообразием функций, которые выполняют эти молекулы в мембранах.
Главные функции липидов мембран состоят в том, что они:
-формируют липидный бислой - структурную основу мембран;
-обеспечивают необходимую для функционирования мембранных белков среду;
-участвуют в регуляции активности ферментов;
-служат «якорем» для поверхностных белков;
-участвуют в передаче гормональных сигналов.
Изменение структуры липидного бислоя может привести к нарушению функций мембран.
Белки мембран. Белки мембран различаются по своему положению в мембране (рис.5). Мембранные белки, контактирующие с гидрофобной областью липидного бислоя, должны быть амфифильными, т.е. иметь неполярный домен. Амфифильность достигается благодаря тому, что:
-аминокислотные остатки, контактирующие с липидным бислоем, в основном неполярны;
-многие мембранные белки ковалентно связаны с остатками жирных кислот
(ацилированы).
Ацильные остатки жирных кислот, присоединенные к белку, обеспечивают его «заякоревание» в мембране и возможность латеральной диффузии. Кроме того, белки мембран подвергаются таким посттрансляционным модификациям, как гликозилирование и фосфорилирование. Гликозилирование наружной поверхности интегральных белков защищает их от повреждения протеазами межклеточного пространства.
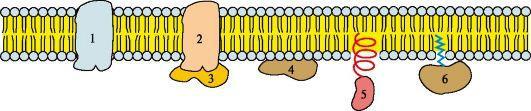
Рис.5 Белки мембран:
1, 2 - интегральные (трансмембранные) белки; 3, 4, 5, 6 - поверхностные белки. В интегральных белках часть полипептидной цепи погружена в липидный слой. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно неполярные аминокислоты. Участки белка, находящиеся в области полярных «головок», обогащены гидрофильными аминокислотными остатками. Поверхностные белки разными способами прикрепляются к мембране: 3 - связанные с интегральными белками; 4 - присоединенные к полярным «головкам» липидного слоя; 5 - «заякоренные» в мембране с помощью короткого гидрофобного концевого домена; 6 - «заякоренные» в мембране с помощью ковалентно связанного ацильного остатка
Наружный и внутренний слои одной и той же мембраны различаются по составу липидов и белков. Эта особенность в строении мембран называется трансмембранней асимметрией.
Белки мембран могут участвовать в:
-избирательном транспорте веществ в клетку и из клетки;
-передаче гормональных сигналов;
-образовании «окаймленных ямок», участвующих в эндоцитозе и экзоцитозе;
-иммунологических реакциях;
-качестве ферментов в превращениях веществ;
-организации межклеточных контактов, обеспечивающих образование тканей и органов.
ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНЫ
Одна из главных функций мембран - регуляция переноса веществ в клетку и из клетки, удержание веществ, которые нужны клетке и выведение ненужных. Транспорт ионов, органических молекул через мембраны может проходить по градиенту концентрации - пассивный транспорт и против градиента концентрации
- активный транспорт.
Пассивный транспорт может осуществляться следующими способами (рис.6,7):
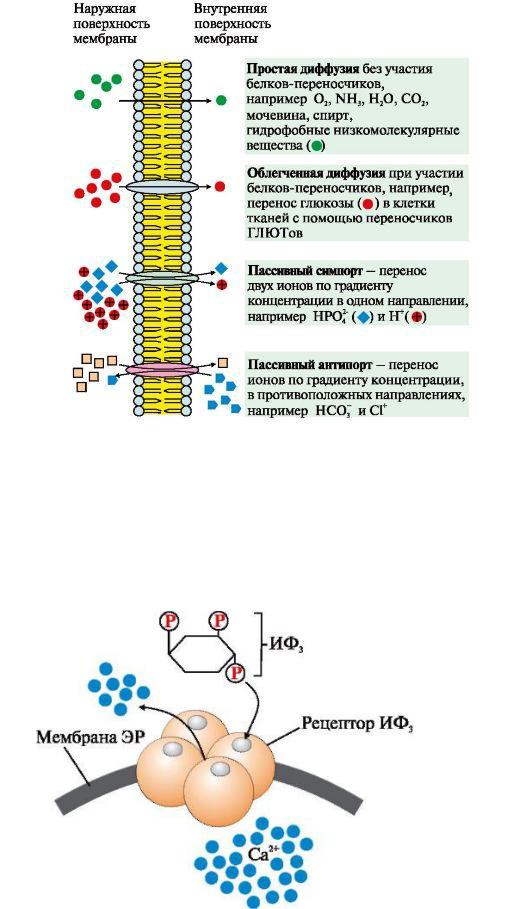
Рис.6 Механизмы переноса веществ через мембраны по градиенту концентрации
К пассивному транспорту относится диффузия ионов по белковым каналам, например диффузия Н+, Са2+, Nа+, К+. Функционирование большинства каналов регулируется специфическими лигандами или изменением трансмембранного потенциала.
Рис.7.Са2+-канал мембраны эндоплазматического ретикулума, регулируемый инозитол-1,4,5-трифосфатом (ИФ3).
ИФ3 (инозитол-1,4,5-трифосфат) образуется при гидролизе мембранного липида ФИФ2 (фосфатидилинозитол-4,5-бисфосфата) под действием фермента фосфолипазы С. ИФ3 связывается специфическими центрами протомеров Са2+- канала мембраны эндоплазматического ретикулума. Изменяется конформация белка и канал открывается - Са2+ поступает в цитозоль клетки по градиенту концентрации
Активный транспорт. Первично-активный транспорт происходит против градиента концентрации с затратой энергии АТФ при участии транспортных АТФаз, например Na+, К+-АТФаза, Н+-АТФаза, Са2+-АТФаза (рис.8). Н+-АТФазы функционируют как протонные насосы, с помощью которых создается кислая среда в лизосомах клетки. С помощью Са2+-АТФазы цитоплазматической мембраны и мембраны эндоплазматического ретикулума поддерживается низкая концентрация кальция в цитозоле клетки и создается внутриклеточное депо Са2+ в митохондриях и эндоплазматическом ретикулуме.
Вторично-активный транспорт происходит за счет градиента концентрации одного из переносимых веществ (рис. 9), который создается чаще всего Na+, К+- АТФазой, функционирующей с затратой АТФ.
Присоединение в активный центр белка-переносчика вещества, концентрация которого выше, изменяет его конформацию и увеличивает сродство к соединению, которое проходит в клетку против градиента концентрации. Вторично-активный транспорт бывает двух типов: активный симпорт и антипорт.
Перенос макромолекул и частиц с участием мембран - эндоцитоз и экзоцитоз.
Перенос из внеклеточной среды в клетку макромолекул, например белков, нуклеиновых кислот, полисахаридов или еще более крупных частиц, происходит путем эндоцитоза. Связывание веществ или высокомолекулярных комплексов происходит в определенных участках плазматической мембраны, которые называются окаймленными ямками. Эндоцитоз, происходящий с участием рецепторов, встроенных в окаймленные ямки, позволяет клеткам поглощать специфические вещества и называется рецептор-зависимым эндоцитозом.
Макромолекулы, например пептидные гормоны, пищеварительные ферменты, белки внеклеточного матрикса, липопротеиновые комплексы, секретируются в кровь или межклеточное пространство путем экзоцитоза. Этот способ транспорта позволяет выводить из клетки вещества, которые накапливаются в секреторных гранулах. В большинстве случаев экзоцитоз регулируется путем изменения концентрации ионов кальция в цитоплазме клеток.
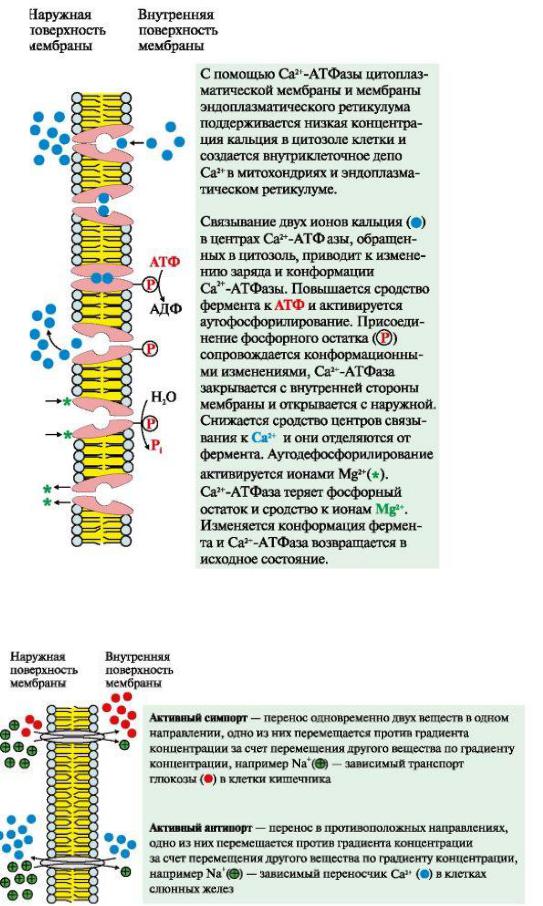
Рис.8 Механизм функционирования Са2+-АТФазы
Рис.9 Вторично-активный транспорт
ТЕМА 4.3. ТРАНСМЕМБРАННАЯ ПЕРЕДАЧА СИГНАЛОВ
Важное свойство мембран - способность воспринимать и передавать внутрь клетки сигналы из окружающей среды. Восприятие клетками внешних сигналов происходит при их взаимодействии с рецепторами, расположенными в мембране клеток-мишеней. Рецепторы, присоединяя сигнальную молекулу, активируют внутриклеточные пути передачи информации, это приводит к изменению скорости различных метаболических процессов.
Сигнальная молекула, специфически взаимодействующая с мембранным рецептором, называется первичным мессенджером. В качестве первичных мессенджеров выступают различные химические соединения - гормоны, нейромедиаторы, эйкозаноиды, ростовые факторы или физические факторы, например квант света. Рецепторы клеточной мембраны, активированные первичными мессенджерами, передают полученную информацию системе белков и ферментов, которые образуют каскад передачи сигнала, обеспечивающий усиление сигнала в несколько сот раз. Время ответа клетки, заключающееся в активации или инактивации метаболических процессов, мышечного сокращения, транспорта веществ из клеток-мишеней, может составлять несколько минут.
Мембранные рецепторы подразделяются на:
-рецепторы, содержащие субъединицу, связывающую первичный мессенджер, и ионный канал;
-рецепторы, способные проявлять каталитическую активность;
-рецепторы, с помощью G-белков активирующие образование вторичных
(внутриклеточных) мессенджеров, передающих сигнал специфическим белкам и ферментам цитозоля (рис.10).
Вторичные мессенджеры имеют небольшую молекулярную массу, с высокой скоростью диффундируют в цитозоле клетки, изменяют активность соответствующих белков, а затем быстро расщепляются или удаляются из цитозоля.