

ФАД-зависимые дегидрогеназы

Убихинон (КоQ)
(КоQ)



Цитохромная система
b c1 c a1 a3
a1 a3
Цитохромоксидаза



Роль дыхательной цепи
•снабжение клетки полезной формой энергии (за сутки в организме в среднем синтезируется до 62 кг АТФ) – превращение окислительной энергии в энергию макроэргических связей
•терморегуляторная функция (при угрозе переохлаждения окисление и фосфорилирование разобщаются).

 Принцип работы дыхательной цепи
Принцип работы дыхательной цепи
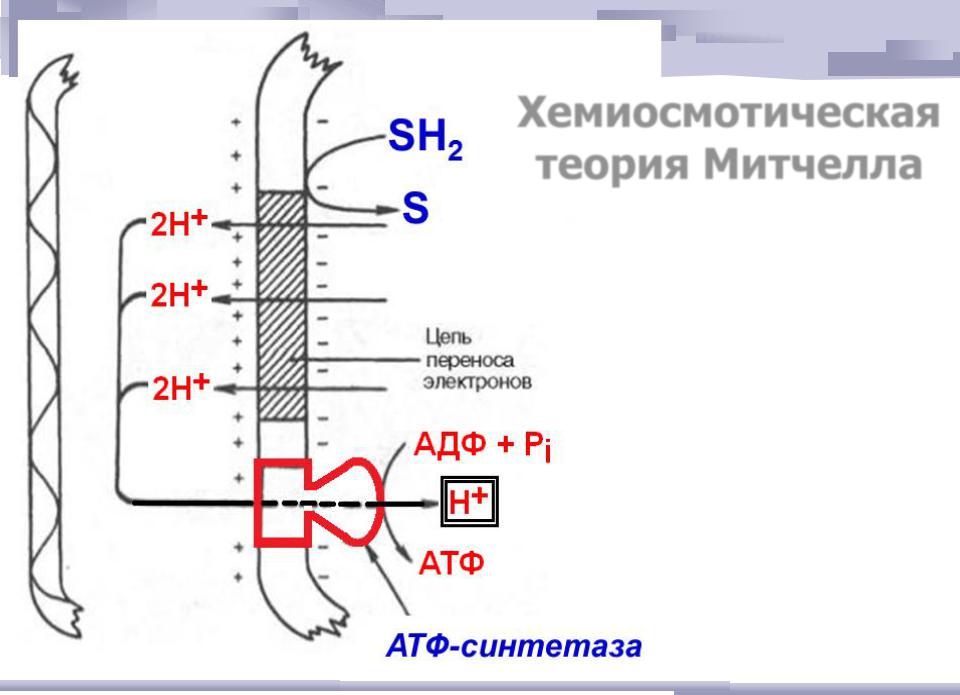
Хемиосмотическая теория Митчелла
Синтез АТФ в дыхательной цепи описывается теорией П. Митчелла: через мембрану митохондрии могут свободно проникать только небольшие незаряженные молекулы, а также гидрофобные молекулы. Энергия, которая выделяется при переносе электронов по цепи переноса электронов, приводит к переносу протонов (Н+) из матрикса митохондрии в межмембранное пространство. Т.е. при движении электронов по дыхательной цепи протоны выбрасываются в межмембранное пространство. Поэтому на внутренней мембране митохондрий образуется градиент концентраций протонов: в межмембранном пространстве Н+ становится много, а в матриксе остается мало. Образуется разность потенциалов 0,14В — наружная часть мембраны заряжена положительно, а внутренняя — отрицательно. Накопившиеся в межмембранном пространстве Н+ стремятся выйти обратно в матрикс по градиенту их концентраций, но митохондриальная мембрана для них непроницаема. Единственный обратный путь в матрикс для протонов — через протонный канал фермента АТФ-синтетазы, которая интегрирован во внутреннюю мембрану митохондрий. При движении протонов по этому каналу в матрикс их энергия используется АТФ-синтазой для синтеза АТФ.

Коэффициент 
 фосфорилирования –
фосфорилирования –
количество фосфатных групп, утилизированных при восстановлении одного атома кислорода, т.е. количество образующихся молекул АТФ при переносе пары электронов и пары протонов по дыхательной цепи
Количество молекул АТФ, а, значит, коэффициент фосфорилирования зависит от того, каким был первичный акцептор водорода: если окисление субстрата происходило при участии НАД-зависимой дегидрогеназы, образуется 3 АТФ (полная дыхательная цепь), если водород акцептировался ФАД-зависимо дегидрогеназой, то первый пункт фосфорилирования отсутствует, и образуется 2 АТФ, а цепь является укороченной. Тип дегидрогеназы зависит от еѐ субстратной специфичности.