

МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ РФ ИРКУТСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
кафедра нормальной физиологии
_________________________________________________
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Учебное пособие для студентов стоматологического факультета
Иркутск, 2009
Печатается по решению Центрального методического координационного совета Иркутского государственного медицинского университета.
Составители: зав. кафедрой нормальной физиологии, д.м.н. профессор Корытов Л.И., доцент кафедры нормальной физиологии к.м.н. Талалаева В.А., ассистент Сидоров А.В.
Рецензенты: зав. кафедрой физиологии человека и животных ИГУ, д.б.н. профессор Гутник И.Н.,
зав. кафедрой патологии с курсом клинической иммунологии и аллергологии ИГМУ, д.м.н. профессор Семинский И.Ж.
2
СОДЕРЖАНИЕ |
|
Общая физиология центральной нервной системы |
4 |
Функции спинного мозга |
11 |
Продолговатый мозг |
14 |
Тонические рефлексы |
18 |
Ретикулярная формация |
20 |
Средний мозг |
21 |
Физиология мозжечка |
24 |
Физиология промежуточного мозга |
27 |
Гипоталамо-гипофизарная система |
31 |
Кора больших полушарий мозга |
32 |
Электрическая активность коры |
39 |
Нервная регуляция вегетативных функций |
40 |
Тесты |
44 |
3
Общая физиология центральной нервной системы
Функции ЦНС
Центральную нервную систему составляют спинной и головной мозг. Основными функциями центральной нервной системы являются:
1) регуляция деятельности всех органов и систем организма и объединение их в единое целое;
2)обеспечение приспособления организма к условиям внешней среды.
Учеловека (и животных с достаточно развитой нервной системой) возможен не только ответ на текущие раздражители, но и интегральный ответ на сложные совокупности раздражителей, а также прогнозирование ситуации и упреждающие воздействия раздражителей реакции, формирующие целенаправленное поведение соответственно потребностям организма.
Основными структурными элементами нервной системы являются нейроны. Нейроны – это клетки способные принимать, обрабатывать, хранить и передавать информацию. Нейроны обычно окружены глиальными клетками, которые не участвуют напрямую в передаче нервных импульсов, однако они выполняют защитную, опорную, изолирующую функции, регулируют состав (ионы, метаболиты, трофические в-ва) интерстициальной жидкости мозга.
Свойства нейронов:
1.Аксонный транспорт (аксоток) - непрерывное движение белков, синтезированных в клетке, синаптических медиаторных веществ и низкомолекулярных факторов, а также органелл из тела нейронов по аксону в область нервных окончаний. Это универсальное явление, наблюдающееся у всех видов животных, имеющих сформированную нервную систему и характерен для любых аксонов. Открыт в 1944-45 г. П. Вейсом. Различают быстрый (200-400 мм/сут) и медленный (0,5-5,0 мм/сут), прямой и ретроградный (обратный). Аксоток регулирует прямые и обратные связи между центром и периферией. Аналогичное явление наблюдается и в дендритах - дендритный ток (дендроток).
2.Генераторная функция - способность к генерации потенциалов. Потенциал действия генерируется в низкопороговой (и соответственно наиболее возбудимой) зоне аксонного холмика и начального сегмента аксона нервной клетки, и проводиться вдоль по аксону к другой нервной клетке или рабочему органу. Процессы, происходящие в активном нейроне, можно представить в виде следующей цепи: потенциал действия в пресинаптическом окончании предыдущего нейрона → выделение медиатора в синаптическую щель → увеличение проницаемости постсинаптической мембраны → ее деполяризация (ВПСП) или гиперполяризация (ТПСП) → взаимодействие ВПСП и ТПСП на мембране сомы и дендритов нейрона → сдвиг мембранного потенциала в случае преобладания возбуждающих влияний → достижение критического уровня деполяризации → возникновение потенциала
4
действия в низкопороговой зоне (мембране аксонного холмика и начального сегмента аксона) нейрона → распространение потенциала действия вдоль по аксону (процесс проведения нервного импульса) → выделение медиатора в окончаниях аксона (передача нервного процесса на следующий нейрон или на рабочий орган).
3.Трофическая функция - способность модулировать, регулировать метаболизм и геном клетки-мишени (например, мышечной клетки).
4.Суммация импульсов - способность суммировать различные влияния, поступающие на нервную клетку. Благодаря суммации нейрон выполняет интегративную функцию, заключающуюся в суммации ВПСП и ТПСП, возникающих в различных участках мембраны сомы и дендритов.
В ответ на одиночный афферентный импульс, идущий к нейрону, в пресинаптической мембране нейрона обычно возникает ВПСП подпороговой величины - 0,1 - 0,2 мв, недостаточной для вызова ответной реакции клетки. Чтобы величина ВПСП достигла критического уровня (порога возникновения ПД - примерно 10 мв), требуется суммация на мембране клетки многих подпороговых ВВСП. Различают пространственную и временную суммацию.
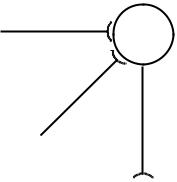
Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам (рис. 1). Одновременное возбуждение синапсов в различных участках мембраны нейрона повышает амплитуду суммарного ВПСП до пороговой величины, возникает ПД. Так, при синхронном раздражении трех афферентных нервов – волокон от короткой малоберцовой мышцы, длинного разгибателя пальцев и длинной малоберцовой мышцы - в мотонейроне длинной малоберцовой мышцы регистрируется ВПСП, равный сумме отдельных ВПСП, получаемых при изолированном раздражении каждого волокна.
///
///
/
Рис. 1. Пространственная суммация возбуждений в ЦНС. На два (или более) синапса одновременно приходит несколько импульсов.
Обычно требуется одновременная активация 50-100 афферентных волокон для получения ответа двигательной клетки спинного мозга.
Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражений (рис.2). Если интервалы между
5
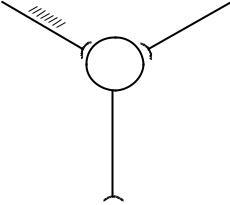
поступающими импульсами достаточно коротки и ВПСП нейрона от предыдущего раздражения не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны не достигнет критического уровня для возникновения ПД.
/
Рис. 2. Временная суммация возбуждений в ЦНС. На один синапс за короткий промежуток времени приходит серия импульсов.
Явления суммации могут наблюдаться при осуществлении отдельных рефлексов (например, рефлексов чихания и кашля в ответ на слабые раздражения слизистой оболочки дыхательных путей).
5.Трансформация ритма возбуждения - способность изменять ритм приходящих к нейронам импульсов; нервные клетки обладают свойством изменять частоту передающихся импульсов. Характер ответного разряда зависит, во-первых, от свойств раздражителя и, во-вторых, от функционального состояния нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях чем сильнее раздражение, тем чаще проявляется ответная импульсация.
При высокой возбудимости нейрона может возникать учащение импульсации (мультипликация ритма). В этом случае афферентная волна вызывает длительные надпороговые ВПСП в клетке, и на каждое одиночное раздражение клетка отвечает залпом из нескольких ПД (пока ВПСП превышает пороговый уровень). При низком уровне возбудимости и лабильности нейрона чаще происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения ПД. В результате на несколько приходящих импульсов нейрон отвечает лишь одним импульсом.
В некоторых крупных клетках, например мотонейронах спинного мозга, имеются специальные механизмы, регулирующие ритм ответных реакций. После каждого ПД в этих клетках наблюдаются выраженные следовые процессы - следовая гиперполяризация мембраны. В этот момент возбудимость клетки понижена. Следующий разряд возникает в клетке лишь тогда, когда заканчивается этот следовой потенциал. Поэтому ритм разрядов мотонейрона, направляющихся к скелетной мышце, обычно не превышает 60 имп/сек. Мелкие клетки, например вставочные нейроны,
6
такого механизма не имеют и могут давать в начальные моменты ответа очень высокую частоту разрядов-до 1600 имп/сек., которая, однако, быстро затухает.
6. Посттетаническая потенциация – работа синапса некоторое время после завершения ритмической стимуляции нейрона (явление, связанное с накоплением ионов Са2+ в пресинаптическом окончании, в результате чего каждый пресинаптический потенциал вызывает высвобождение большего числа квантов медиатора). С свойством посттетанической потенциации связано явление последействия - рефлекторные акты заканчиваются не одновременно с прекращением действия раздражителя, а через некоторый период (его связывают также с реверберацией).
7.Утомление - связано с нарушением передачи возбуждения в межнейронных синапсах;
8. Лабильность - это скорость протекания элементарных реакций, лежащих в основе его возбуждения. Наиболее удобная мера лабильности нейрона – максимальная частота импульсов, воспроизводимых нейроном без трансформации в единицу времени. Разные по функциям и величине нервные клетки обладают различными величинами лабильности. Даже в пределах одного нейрона лабильность разных его структур (дендритов, сомы, начального сегмента и аксона) резко различается.
Величина лабильности нейрона определяется уровнем его мембранного потенциала. Лишь при определенной величине мембранного потенциала достигается оптимальный уровень возбудимости и лабильности нервной клетки, а также наиболее высокий уровень ее ритмической активности, что является важным условием для передачи информации в нервной системе и осуществления целесообразных реакций. Например, наибольший максимальный ритм мотонейронов спинного мозга (до 100 импульсов в 1 сек.) отмечается при средней, оптимальной, величине мембранного потенциала-55 мв (т. е. при повышенной возбудимости мотонейрона).
В естественных условиях деятельности частота разрядов мотонейрона обычно не превышает 50-60 импульсов в 1 сек. Лишь при мощных влияниях вышележащих отделов нервной системы резко изменяется мембранный потенциал мотонейронов и максимальная частота их разрядов может повыситься до 100 и даже до 300 импульсов в 1 сек. Однако разряды с такой частотой очень кратковременны. Они сменяются более медленным и устойчивым ритмом активности. В мелких промежуточных нейронах спинного мозга частота разрядов в начальные моменты активации достигает 600800 импульсов в 1 сек., а иногда превышает 1000 импульсов в 1 сек. Такая частота необходима для резкого и быстрого воздействия этих клеток на мотонейроны; через некоторое время она резко замедляется.
Нейроны объединены в специфически организованные нейронные цепи и центры, составляющие различные функциональные системы мозга. Нервным центром называют совокупность (ансамбли) нервных клеток, необходимых для осуществления
7
какой-либо функции. Сложные реакции в целостном организме обычно осуществляются с участием многих нервных центров, расположенных в различных этажах центральной нервной системы, связанных в нервные цепи. Например, произвольное изменение дыхания осуществляется человеком при участии центров коры больших полушарий, особого (пневмотаксического) центра в мосте, дыхательного центра продолговатого мозга и центров спинного мозга, иннервирующих дыхательные мышцы. Для выявления функций нервного центра используют ряд методов: метод раздражения (усиление его деятельности), метод разрушения или удаления –
перерезка или экстирпация (нарушение регулируемой центром функции), электрофизиологические методы - регистрация электрических явлений в нервном центре с помощью микро- и макроэлектродной техники и др. Так, функциональное значение, а также локализация дыхательного центра в продолговатом мозгу были выявлены путем последовательных перерезок головного мозга. При перерезках выше области его расположения (выше продолговатого мозга) дыхание сохранялось, а при перерезках ниже продолговатого мозга дыхательные движения исчезали.
Принципы распространения возбуждения в ЦНС 1. Дивергенция – поступление афферентной информации одновременно к разным
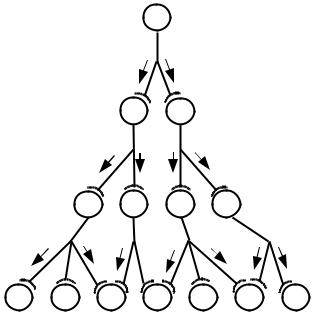
(многим) участкам нервной системы. Дивергенция пути обеспечивает расширение области действия стимула, т. е. иррадиацию возбуждения и торможения. Она осуществляется благодаря многочисленным взаимосвязям нейронов одной рефлекторной дуги с нейронами других рефлекторных дуг, так что при раздражении одного рецептора возбуждение в принципе может распространяться в центральной нервной системе в любом направлении и на любую нервную клетку (рис. 3).
Рис. 3. Дивергенция (иррадиация) возбуждения в ЦНС
8
Чем сильнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Это явление можно наблюдать на спинальной (перерезка между спинным и головным мозгом) лягушке. Слабое давление на пальцы задней лапки вызывает ответный рефлекс сгибания этой же лапки. Небольшое усиление давления приводит к сгибанию другой задней лапки, хотя рецепторы последней не раздражаются. При еще более сильном раздражении волна возбуждения охватывает вышележащие и нижележащие нервные центры и вызывает движения верхних конечностей (сначала на стороне тела, подвергшейся раздражению, а затем на противоположной).
Этот ответ возникает в результате того, что в сферу возбуждения помимо нервных центров одноименной половины спинного мозга вовлекаются центры другой его половины.
Аналогичное явление иррадиации возбуждения можно наблюдать при действии различных раздражении в коре больших полушарий.
Процесс иррадиации играет положительную роль при формировании новых реакций организма (ориентировочных реакций, условных рефлексов). Активация большого количества различных нервных центров позволяет отобрать из их числа наиболее нужные для последующей деятельности, т. е. совершенствовать ответные действия организма. Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные связи - условные рефлексы.
Вместе с тем иррадиация возбуждения может оказать и отрицательное воздействие на состояние и поведение организма. Так, иррадиация возбуждения в центральной нервной системе нарушает тонкие взаимоотношения, сложившиеся между процессами возбуждения и торможения в нервных центрах, и приводит к расстройству двигательной деятельности.
2. Конвергенция - приходящие импульсы с периферии в ЦНС по афферентным волокнам, могут сходиться (конвергировать) к одним и тем же промежуточным и эффекторным нейронам (рис. 4).
В низших отделах нервной системы – спинном и продолговатом мозгу - конвергенция выражена гораздо меньше. Нейроны этих отделов получают информацию от рецепторов сравнительно небольших участков тела - рецептивных полей одного и того же рефлекса. В надсегментарных отделах, особенно в коре больших полушарий, происходит конвергенция импульсов различного происхождения от разных рефлекторных путей. Нейроны надсегментарных отделов могут получать
9

сигналы о световых, звуковых, проприоцептивных и прочих раздражениях, т.е. сигналы разной модальности.
Рис. 4. Конвергенция возбуждения в центральной нервной системе
На теле нейронов постоянно изменяются «конвергентные узоры» - возбужденные и заторможенные участки. Подсчитано, что размеры рецептивных полей корковых нейронов, т.е. участков тела, от которых к ним могут поступать афферентные раздражения, в 16100 раз больше, чем размеры тех же полей для афферентных клеток спинальных рефлекторных дуг. Благодаря такому разнообразию поступающей информации в нейронах вышележащих отделов головного мозга может происходить ее широкое взаимодействие, сопоставление, отбор, выработка адекватных реакций и установление новых связей между рефлексами.
4. Реверберация - циркуляция возбуждения по замкнутым нейронным цепям.
Одной из основных функций центральной нервной системы является интегративная и координационная деятельность. В основе последней также лежат некоторые принципы:
1. Принцип реципрокности - взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов (у однонаправленных рефлексов это взаимное усиление, у противоположных рефлексов - взаимное торможение).
3. Окклюзия – если при раздельной стимуляции каждого из двух входов к нейронной популяции возбуждение является надпороговым во всех нейронах, то одновременная стимуляция обоих входов вызовет надпороговое возбуждение в меньшем числе нейронов, чем при раздельной активации входов, так, что общее число
10
возбужденных нейронов не достигнет алгебраической суммы нейронов, возбужденной при раздельной активации.
4. Принцип обратной связи - связь выхода системы с ее входом через усилительное звено с положительным коэффициентом усиления (положительная обратная связь) или с отрицательным усилением (отрицательная обратная связь).
5.Принцип общего "конечного пути" - английским физиологом Ч. Шерингтоном введено понятие о мотонейроне как общем конечном пути двигательной системы, т.е. клетке, которая производит сопоставление между возбуждающими и тормозящими влияниями (число синапсов на мотонейроне может достигать 20 тыс.);
6.Доминанта - общий рабочий принцип нервной системы; под этим термином понимают господствующий очаг возбуждения, направляющий работу центральной нервной системы в каждый данный момент; может возникать на любых этажах нервной системы. Характеризуется повышенной возбудимостью, стойкостью, способностью к суммированию и инерцией возбуждения (т.е. способностью продолжать реакцию, когда стимул уже миновал); подкрепляться посторонними импульсами независимо от их характера и тормозить другие импульсы.
Функции спинного мозга.
Рефлекторная функция СМ:
Спинной мозг является филогенетически самым древним отделом ЦНС. В нем располагаются нейроны нескольких типов. Около трех процентов составляют двигательные нейроны или мотонейроны. Они, в свою очередь подразделяются на
альфамотонейроны фазические (быстрые) и альфамотонейроны тонические (медленные), а также гаммамотонейроны. Кроме того, более 95% приходится на вставочные или интернейроны, среди которых выделяют собственные спинальные и проекционные.
В структурах спинного мозга замыкается большое количество рефлекторных дуг, принимающих участие в регуляции соматических и вегетативных функций организма.
Часть спинномозговых рефлексов связана с деятельностью нейронных механизмов самого спинного мозга (сегментарные рефлексы), другая связана с деятельностью различных центров головного мозга (надсегментарные рефлексы) и поэтому рефлексы спинного мозга могут отличаться самой различной степенью сложности.
Существует несколько классификаций спиномозговых рефлексов, основные из них две следующие:
1-я - по рецепторам, раздражение которых вызывает рефлекс. По ней различают:
Проприорецептивные, висцерорецептивные и кожные рефлексы.
Проприорецептивные - запускаются рецепторами мышц, суставов и сухожильными рецепторами. Отсюда их первоначальное название - сухожильные рефлексы:
11
Обнаружены немецкими неврологами В.Эрбом и К.Вестфалем в 1875 г. и с тех пор широко применяются в клинике как тест при исследованиях рефлекторной возбудимости спинного мозга человека.
различают две формы сухожильных рефлексов:
- фазический рефлекс на растяжение (возникает в ответ на очень кратковременное растяжение мышцы напр. удар); именно они были описаны В.Эрбом и К.Вестфалем в 1875 г.: например, коленный рефлекс.
- тонический рефлекс на растяжение (возникает в ответ на длительное растяжение, продолжающееся десятки сек.); (открыт в 1924 г. Ч.Шеррингтоном). Основным отличием от фазических рефлексов являются временные характеристики их течения: фазический рефлекс представляет собой кратковременную (фазную реакцию), рефлексы же растяжения носят длительный, тонический характер (растяжение мышцы - лучше экстензора - вызывает рефлекторное тоническое сокращение мышечных волокон, которое противодействует растяжению); Такие рефлексы ещё называются миотатическими, а фазические просто сухожильными. Подобного рода рефлексы используются в организме для поддержания позы и регуляции движений).
Висцерорецептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов), могут быть соматическими и вегетативными.
Соматические проявляются в сокращении мышц передней брюшной стенки, грудной клетки и разгибателей спины.
Вегетативные осуществляются при участии преганглионарных нейронов вегетативной нервной системы (подразделяются по её отделам):
-рефлексы симпатической нервной системы (сосудодвигательные, железистые и т.д.);
-рефлексы парасимпатической нервной системы (регуляция сокращений гладкой мускулатуры мочевого пузыря, кишечника и т.д.).
Последняя группа - кожные рефлексы являются защитными.
2-я классификация обьединяет спинальные рефлексы по органам - эффекторам рефлекса: (согласно ей выделяют 3 группы) рефлексы конечностей, брюшные, органов таза.
Рефлексы конечностей - наиболее обширная группа рефлексов, включает:
-сгибательные рефлексы - представляют собой четко дифференцированные, мощные рефлекторные реакции, которые хорошо развиты у всех наземных позвоночных животных. Все сгибательные рефлексы имеют сходное функциональное значение - это рефлексы защитного типа, направленные на удаление животного от сильных повреждающих воздействий или на сбрасывание с поверхности тела источников таких раздражений. Рефлекторная дуга этих рефлексов может иметь полисинаптический характер, проявляются при раздражении болевых рецепторов кожи, мышц и внутренних органов.
Например, погружение в слаб раствор серной кислоты лапки лягушки в опыте Тюрка или щипок конечности пинцетом вызывает отдёргивание лапки ...
12