

111. Строение, физиологические свойства и функции гладкомышечных клеток. Виды хеморецепторов мембраны гладкомышечных клеток.
Физиологические свойства гладких мышц:
- возбудимость (ниже, чем в нервном волокне, что объясняется низкой величиной мембранного потенциала)
- низкая проводимость, порядка 10–13 м/с
- рефрактерность (занимает по времени больший отрезок, чем у нервного волокна)
- лабильность
- сократимость (способность укорачиваться или развивать напряжение)
- нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного частичного сокращения – тонуса
- самопроизвольную автоматическую активность
- сокращение в ответ на растяжение
- пластичность (уменьшение растяжения при увеличении растяжения)
- высокую чувствительность к химическим веществам
Физиологические функции гладких мышц:
- поддержание давления в полых органах
- регуляция давления в кровеносных сосудах
- опорожнение полых органов и продвижение их содержимого
Виды хеморецепторов мембраны гладкомышечных клеток.
1) альфа- и бета- адренергические: Взаимодействие адреналина (норадреналина) с этими рецепторами может вызвать различные эффекты. Активизация а-рецепторов приводит к снижению уровня цАМФ в середине клетки, увеличение концентрации Са2 +, сокращение мышц и сужения сосуда. Взаимодействие с В-рецепторами сопровождается увеличением уровня цАМФ, снижением концентрации свободного Са2 + и расслаблением гладкомышечной клетки.
2) холинергические: Ацетилхолин через М-холинорецепторы приводит к увеличению уровня цГМФ. Вследствие этого снижается уровень свободного внутриклеточного Са2 + и клетка расслабляется.
3) гистаминергические: Относительно гистамина есть два типа рецепторов – Н1 и Н2. Активизация H1-рецепторов сопровождается деполяризацией мембраны, увеличивая выход К + из клетки. При этом снижается сила сокращения мышцы, но сокращение сохраняется, потому что при активизации Н2-рецепторов уменьшается выход К + из клетки и Са2 + из депо в цитоплазму, что приводит к расслаблению гладких мышц.
112. Виды межклеточных контактов. Классификация синапсов. Строение электрических и химических синапсов.
Контакты между двумя клетками
Плотные контакты
Септированные контакты
Адгезивные контакты
Щелевые контакты
Десмосомы
Плазмодесмы

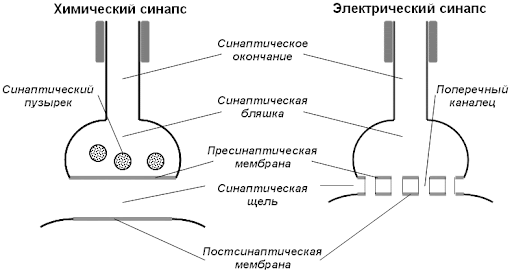
113. Механизм проведения возбуждения в электрических и химических синапсах нервной системы. Постсинаптические потенциалы в нервных синапсах, их природа.
Механизм проведения возбуждения в электрических синапсах
Две соседние клетки прилегают друг к другу так тесно, что сопротивление двух их мембран протекающему через них электрическому току сравнимо с сопротивлением остальной, внесинаптической области мембраны. При возбуждении 1-ой клетки натриевый ток (INa) входит в нее через открытые Nа-каналы и выходит через пока невозбужденные участки мембраны; при этом часть тока входит через участок мембранного контакта во 2-ю клетку, вызывая ее деполяризацию. Разумеется, здесь уровень деполяризации гораздо ниже - скажем, в 10 раз, чем в 1-ой клетке, однако он может оказаться выше порога генерирования потенциала действия во 2-ой клетке. Часто такая деполяризация подпороговая, и тогда 2-ая клетка возбуждается только в результате суммации синаптических потенциалов, возникающих в результате химической или электрической передачи от других клеток.
Большинство известных электрических синапсов образованы большими пресинаптическими аксонами, контактирующими со сравнительно мелкими волокнами постсинаптических клеток. Передача информации в них происходит без химического посредника, а между взаимодействующими клетками очень небольшое расстояние: ширина синаптической щели около 3,5 нм, тогда как в химических синапсах она варьирует от 20 до 40 нм. Кроме того, синаптическую щель пересекают соединительные мостики – специализированные белковые структуры, образующие т.н. коннексоны (от англ. connexion – соединение).
Механизм проведения возбуждения в химических синапсах нервной системы.
Передача в синапсе имеет два главных этапа.
1. Преобразование электрического сигнала в химический (электросекреторное сопряжение). Потенциал действия (ПД), поступивший в пресинаптическое окончание, вызывает деполяризацию его мембраны, открывающую потенциалзависимые Са-каналы. Ионы кальция входят, согласно концентрационному и электрическому градиентам, внутрь клетки, что ведет к увеличению его содержания в цитозоле в 10—100 раз. Ионы кальция активируют фосфорилирование синаптосина, что ослабляет связь везикулы с цитоскелетом, и везикула перемещается вдоль микротрубочек на позицию у активной зоны. При контакте везикулы с пресинаптической мембраной происходит ферментативное «плавление» ее стенки, а также активация белка синаптопорина, формирующего канал, через который медиатор выходит в синаптическую щель посредством первично-активного транспорта — экзоцитоза.
Молекулы медиатора, поступившие в синаптическую щель, диффундируют к постсинаптической мембране и вступают во взаимодействие с ее рецепторами. Открывание каналов в химических синапсах происходит в результате связывания медиатора или его агониста с комплексом рецептор-канал.
Удаление медиатора осуществляется путем его диффузии из щели в окружающую жидкость и разрушения его под действием ацетилхолинэстеразы. При этом большая часть (около 60 %) холина захватывается обратно пресинаптическим окончанием. Значительная доля высвобожденного ацетилхолина разрушается уже в ходе диффузии через синаптическую щель, не успевая достигнуть рецепторов, и через несколько миллисекунд его практически не остается: синапс вновь готов к передаче возбуждения.
2. Преобразование химического сигнала обратно в электрический. Этот этап осуществляется в постсинаптической мембране. Действие молекул медиатора на ее рецепторы ведет к открытию ионных каналов и перемещению ионов, имеющих высокий электрохимический градиент на протяжении канала. Канал хорошо проницаем для Nа+ и К+, плохо проницаем для Са2+. Входящий в клетку ток натрия резко преобладает над выходящим из клетки током калия, так как ион Nа+ движется в клетку согласно концентрационному и электрическому градиенту (клетка внутри имеет положительный заряд), а ион К+ выходит из клетки только согласно концентрационному градиенту, причем вопреки электрическому (снаружи клетка имеет положительный заряд). Поэтому суммарный ток ионов Nа+ в клетку превосходит ток К+ из клетки, что и приводит к деполяризации постсинаптической мембраны (концевой пластинки). Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП), который в нервно-мышечном синапсе называют потенциалом коцевой пластинки (ПКП).
Постсинаптические потенциалы в нервных синапсах (ВПСП, ТПСП), их природа.
В результате открывания каналов изменяется потенциал субсинаптической мембраны. Такое изменение называется постсинаптическим потенциалом.
В ЦНС возбуждающими являются холин-, адрен-, дофамин-, серотонинергические синапсы и некоторые другие. При взаимодействии их медиаторов с соответствующими рецепторами, открываются хемозависимые натриевые каналы. Ионы натрия входят в клетку через субсинаптическую мембрану. Происходит ее местная или распространяющаяся деполяризация. Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП).
Тормозными являются глицин- и ГАМК-ергические синапсы. При связывании медиатора с хеморецепторами, активируются калиевые или хлорные хемозависимые каналы. В результате ионы калия выходят из клетки через мембрану.
Ионы хлора входят через нее. Возникает только местная гиперполяризация субсинаптической мембраны. Она называется тормозным постсинаптическим потенциалом (ТПСП).
114. Механизм проведения возбуждения в нервно-мышечных синапсах. Роль Ca2+ в механизме синаптического процесса.
Механизм проведения возбуждения в нервно-мышечных синапсах.
Нервно-мышечные синапсы обеспечивают проведение возбуждения с нервного волокна на мышечное благодаря медиатору ацетилхолину, который при возбуждении нервного окончания переходит в синаптическую щель и действует на концевую пластинку мышечного волокна. В пресинаптической терминали образуется и скапливается в виде пузырьков ацетилхолин. При возбуждении электрическим импульсом, идущим по аксону, пресинаптической части синапса ее мембрана становится проницаемой для ацетилхолина.
Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются ее кальциевые каналы. Ион Са2+ входит в пресинаптическую часть синапса из синаптической щели. Ацетилхолин высвобождается и проникает в синаптическую щель. Здесь он взаимодействует со своими рецепторами постсинаптической мембраны, принадлежащей мышечному волокну. Рецепторы, возбуждаясь, открывают белковый канал, встроенный в липидный слой мембраны. Через открытый канал внутрь мышечной клетки проникают ионы Na+, что приводит к деполяризации мембраны мышечной клетки, в результате развивается так называемый потенциал концевой пластинки (ПКП). Он вызывает генерацию потенциала действия мышечного волокна.
Нервно-мышечный синапс передает возбуждение в одном направлении: от нервного окончания к постсинаптической мембране мышечного волокна, что обусловлено наличием химического звена в механизме нервно-мышечной передачи.
Ca 2+ активирует белки, отвечающие за слияние содержащих медиатор пузырьков с пресинаптической мембраной. 115. Торможение в нервной системе. Виды торможения.Природа пре-, постсинаптического, возвратного и пессимального торможения.
Торможение в нервной системе. Виды торможения.
Торможение — биологический процесс, направленный на ослабление или предотвращение возникновения процесса возбуждения.
Торможение в ЦНС способствует определенной координации выполняемой функции. При этом блокируется деятельность нейронов и центров, которые в данный момент не требуются для выполнения приспособительной реакции. Кроме того, торможение выполняет и защитную функцию, предохраняя нервные клетки от перевозбуждения и истощения при действии сильных раздражителей.
Различают несколько видов торможения в нервной системе.
1. Постсипаптическое торможение развивается в случаях, когда тормозной медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что нервная клетка не может генерировать потенциал действия. Постсипаптическое торможение может быть обусловлено длительной деполяризацией или гиперполяризацией, возникающей в постсинаптической мембране вследствие взаимодействия медиатора с рецепторами, открывающими калиевые и хлорные каналы. Наиболее распространенными тормозными медиаторами являются гамма-аминомасляная кислота и глицин.
А) Возвратное постсинаптическое торможение - это такое торможение, при котором тормозные вставочные нейроны (клетки Реншоу) действуют на те же нервные клетки, которые их иннервируют. Примером возвратного постсинаптического торможения может служить торможение в мотонейронах спинного мозга. Этот вид торможения обеспечивает, например, поочередное сокращение и расслабление скелетных мышц — сгибателей и разгибателей, что необходимо для координации движений конечностей при ходьбе.
Б) Латеральное постсинаптическое торможение обусловлено тем, что тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. В результате в этих соседних клетках развивается очень глубокое торможение, называемое латеральным, так как образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону и инициируется им.
В) Реципрокное торможение, примером которого является торможение нервных центров мышц-антагонистов, заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны. Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей. Если бы возбуждались одновременно центры мышц-сгибателей и мышц- разгибателей, сгибание конечности в суставе было бы невозможно.
2. Пресинаптическое торможение связано с тем, что в пресинаптическом окончании может развиваться продолжительная деполяризация мембраны, которая приводит к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения и импульсы не могут пройти через зону деполяризации. Следовательно, не происходит выделения медиатора в синаптическую щель в достаточном количестве и не возбуждается постсинаптический нейрон. В ЦНС имеется огромное число тормозных нейронов, в частности клетки Реншоу. Эти тормозные нейроны синтезируют специфические тормозные медиаторы и осуществляют реакцию торможения.
Постсинаптическое торможение. Тормозные нейроны. В настоящее время установлено, что в ЦНС наряду с возбуждающими нейронами существуют и особые тормозные нейроны. Примером может служить клетка Реншоу в спинном мозге. Реншоу открыл, что аксоны мотонейронов перед выходом из спинного мозга дают одну или несколько коллатералей, которые заканчиваются на особых клетках, чьи аксоны образуют тормозные синапсы на мотонейронах данного сегмента. Благодаря этому возбуждение, возникающее в мотонейроне, по прямому пути распространяется на периферию к скелетной мышце, а по коллатерали активирует тормозную клетку, которая подавляет дальнейшее возбуждение мотонейрона. Это механизм, автоматически охраняющий нервные клетки от чрезмерного возбуждения. Торможение, осуществляющееся при участии клеток Реншоу, получило название возвратного постсинаптического торможения. Тормозным медиатором у клетки Реншоу является глицин.
Пресинаптическое торможение связано с тем, что в пресинаптическом окончании может развиваться продолжительная деполяризация мембраны, которая приводит к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения и импульсы не могут пройти через зону деполяризации. Следовательно, не происходит выделения медиатора в синаптическую щель в достаточном количестве и не возбуждается постсинаптический нейрон. В ЦНС имеется огромное число тормозных нейронов, в частности клетки Реншоу. Эти тормозные нейроны синтезируют специфические тормозные медиаторы и осуществляют реакцию торможения. Активация тормозного нейрона вызывает деполяризацию мембраны терминалей в афферентных нейронах, что затрудняет процесс проведения потенциала действия. Медиатором в таких аксо-аксональных синапсах служит гамма-аминомасляная кислота или другой тормозной медиатор. Деполяризация является следствием повышения проницаемости мембраны для ионов хлора, в результате эти ионы выходят из клетки.
Возвратным торможением называют нейронную цепь, в которой тормозные вставочные нейроны действуют на те же нервные клетки, которые их возбуждают. Характерный пример возвратного торможения существует в мотонейронах. Мотонейроны посылают коллатерали к вставочным нейронам, аксоны которых образуют тормозные синапсы на мотонейронах. Такая тормозная цепь называется торможением Реншоу, а тормозной вставочный нейрон клеткой Реншоу В такой цепи усиление возбуждения, поступающего к мышце, усиливает торможение мотонейрона под действием клетки Реншоу. Это пример торможения по принципу обратной связи.