

Зрительная кора
Следующий этап анализа зрительных стимулов связан с функцией коры больших полушарий. В каждом полушарии представлены лишь контрлатеральные половины полей зрения обоих глаз.Нейроны, получающие импульсацию от каждого глаза, образуют в области коры своеобразную "чрезполосицу".
В коре нейроны сгруппированы в вертикальные колонки одинакового функционального назначения. В их формировании можно выделить ту особенность, что область коры, получающая информацию от центральной ямки - зоны наивысшей остроты зрения, примерно в 35 раз больше участка такой же величин периферии сетчатки. Хотя эти представления весьма относительны: именно в этих отделах сетчатки плотность рецепторных клеток наибольшая, так что число ганглиозных клеток сетчатки, связанных с одной корковой колонкой нейронов имеет примерно одинаковый диаметр - около 1 мм.
В первичной зоне коры лишь небольшая часть нейронов реагирует на простые стимулы, типа свет или темнота. Другая группа нейронов отвечает на сравнительно простые контуры определенной ориентации, изломы их. Однако большинство нейронов коры обеспечивает опознание сложных и сверхсложных РП. Сложные связаны с РП, стимулом для которых являются, к примеру, границы между темным и светлым причем границы определенной ориентации, или разрывы границ определенной ориентации и ширины и т.п. Сверхсложные РП части нейронов реагируют на границы между светлым и темным строго определенной ориентации и ограниченной длины, определенные углы и т.п.
Кроме этих отделов коры в ней имеется еще ряд нейронных полей, причастных к обработке зрительной сигнализации. Первичная зрительная кора проектируется в большое количество частей мозга, которые принамают участие в обработке зрительной сигнализации. Так, в теменной доле коры (поле 17 и др.) имеются нейроны, осуществляющие дальнейшую (после зрительных зон затылочной доли) обработку информации. Этот отдел играет значительную роль в регуляции движения глаз и головы для фиксации объекта. Нейроны, участвующие в обработке зрительной информации имеются и в других отделах ассоциативных областей коры (височная доля, лобная доля). Но для осознания увиденного требуется «путешествие» возбуждения по различным отделам коры больших полушарий с повторным возвратом его к зрительным зонам коры (6.29).
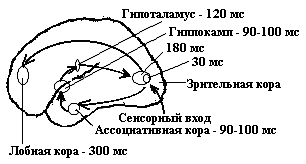
Рис. 6. 29. Схема кольцевого движения возбуждения при возникновении зрительного ощущения и время регистрации его в нервных центрах после предъявление раздражителя (по А.М. Иваницкому).
В результате в коре больших полушарий, например, для восприятия какой-либо буквы одновременно должны возбуждаться многие нейроны, каждый из которых реагирует на определенную часть буквы. Их возбуждение создает своеобразную нейронную мозаику, которая в процессе обучения и позволяет узнавать эту букву как целое. Эти процессы совершаются во вторичной и третичной зрительных областях коры.
6. 12.6. Восприятие цвета
Глаз человека в состоянии различать не только формы, поверхности или оттенки серого цвета. Ему доступны для различения волны в диапазоне от 400 до 760 нм, сочетание которых воспринимаются как различные цвета. Непосредственно примыкающие к ним области инфракрасного и ультрафиолетового света не вызывают никаких цветовых ощущений, хотя при высокой интенсивности они могут ассоциироваться с весьма слабыми (серыми) тонами. Человек может различать до 7 миллионов различных цветовых оттенков. Хроматические оттенки можно охарактеризовать тремя качествами: тоном, насыщенностью и светлотой. Вся гамма цветов - от красного до фиолетового может быть представлена постепенным переходом от одного оттенка к другому.
Каждый цвет имеет свою волновую характеристику. Разновидности оттенков красного цвета занимают часть спектра с длиной волн от 723 до 647 нм, зеленого – 575-492 нм, синего – 492-450 нм. Максимум длины волны чисто красного цвета - 570 нм, зеленого - 536 нм, голубого - 445 нм. Смешение этих цветов дает промежуточные цвета. Указанные три цвета признаны международной конвенцией в качестве главных (первичных) цветов. Равномерное смешивание их дает белый цвет, а, к примеру, смешивание красного и зеленого цветов дает промежуточный желтый цвет. Эти представления легли в основутрехкомпонентной теории цветового зрения (Юнг, Гельмгольц). Полагают, что на уровне рецепторов цветовое зрение обеспечивается наличием в сетчатке как минимум трех типов колбочек, каждая из которых работает как независимые приемники (рис. 6.29). Одни колбочки содержат пигмент, реагирующий на красный цвет, другие - на зеленый, третьи - фиолетовый. Любой цвет оказывает влияние на все три типа колбочек, но чувствительность к "своему" выше. Комбинация возбуждения их обрабатывается по всей афферентной "дорожке", вплоть до коры больших полушарий, и только комплекс возникающих физиологических процессов воспринимается в нашем сознании как соответствующий цвет.
Синтез зеленочувствительного и красночувствительного пигментов зависит от генов, расположенных в Х-хромосоме. Поэтому дальтониками,то есть людьми, не воспинимающими длину волны красного цвета, являются около 8% мужчин и лишь 0,4% - женщин, причем около 2% мужчин - дихроматы. Трихроматное нарушение чувствительности не является половозависимой, так как ген, кодирующий синюючувствительность, не связан с Х-хромосомой.
Теория оппонентных цветов (Геринг). Было замечено, что при рассматривании находящихся рядом некоторых цветов, можно обнаружить появление дополнительного тре-тьего цвета или исчезновение какого-либо цвета. Так, например, серый круг в окружении ярко зеленого кольца видится красным. Геринг, предложивший теорию оппонентных цветов, считал, что имеется четыре основных цвета, на основе которых можно выделить попарные их цветоконтрастные сочетания: зелено-красное и желто-синее. Предлагалось, выделить три типа колбочек: воспринимающих указанные две пары цветовых контрастов и бело-черный. Другие цветовые ощущения рождаются из сочетания указанных трех. Однако предположение о наличие такого типа колбочек в сетчатке не подтвердилось.
В последующем , когда научились отводить биопотенциалы от отдельных рецепторов и нервных клеток, было показано, что верными, по-видимому, являются обе вышеуказанные теории. Трехкомпонентная теория пригодна для описания процессов, происходящих на уровне колбочек. А вот обработка цветовой информации на более высоких уровнях, начиная с ганглиозных нейронных цепей происходит по типуодновременного цветового контраста. Были обнаружены ганглиозные клетки, имеющие рецептивные поля указанных выше цветовых сочетаний. Освещение колбочек одним цветом, возбуждая деполяризующиеся биполярные клетки, возбуждает ганглиозную клетку. Другие цвета ингибируют ганглиозную клетку через гиперполяризующиеся биполярные или горизонтальные клетки. В некоторых случаях спектральная чувствительность периферии РП гангиозных клеток является зеркальным отражением кривых для их центра. Нейроны латеральных коленчатых тел, также представлены по типу цветоконтрастных рецептивных полей.
Хорошим клиническим доказательством наличия цветоспецифических колбочек являются люди, у которых врожденно отсутствуют колбочки, воспринимающие тот или иной цвет. Наиболее часто встречаются дальтоники (протанотропы)- люди, не воспринимающие красный цвет. Дальтонизм встречается у 8 % мужчин и обусловлен генетическим дефектом в непарной Х-хромосо-ме. Протанотропы не воспринимают красный цвет, а сине-голубые лучи им кажутся бесцветными. Значительно реже встречаютсядейтеранопия ("зеленослепые"), когда зеленые цвета не отличаются от темнокрасных и голубых. Еще реже встречаются люди стританопией, которые не воспринимают лучи синего и фиолетового цвета. Встречается и полная цветовая слепота вследствие полного поражения колбочкового аппарата. Такой человек все предметы видит лишь в виде различных оттенках серого цвета.
Однако, цветовые характеристики на уровне сетчатки и подкорковых структур только начинают анализироваться. Окончательные анализ их происходит на уровне коры больших полушарий. Человек, имеющий дефект цветового зрения, в процессе обучения приспосабливается, частично компенсируя свой недостаток.
Для обнаружения нарушения дефекта восприятия цветов можно воспользоваться специальными полихроматическими таблицами.