

6.10.4. Центральные отделы вестибулярной системы
Первые центры вестибулярного анализатора находятся в ядрах стволамозга, куда подходит большинство нервных волокон от рецеторов. Некоторая часть волокон вестибулярных нервов идет к различным образованияммозжечка. Нейроны вестибулярных ядер контактируют с многими отделами ЦНС (рис. 6.11), как то:
- α- и γ-мотонейронами мышц-разгибателей спинного мозга,
- ядрами глазодвигательного нерва,
- мозжечка,
- ретикулярной формации,
- таламус (а отсюда с сенсомоторными отделами коры),
- гипоталамусом (связь с регуляцией функций внутренних органов).
В результате при интенсивном раздражении рецепторов вестибулярного анализатора возникают моторные рефлексы, глазнойнистагм(вначале происходит медленный поворот глазного яблока в сторону, противоположную вращению, а затем быстрое движение в сторону вращения, когда глаз как бы "догоняет" вращающуюся голову),вегетативные расстройства(изменение частоты сердечных сокращений, сужение сосудов кожи, потоотделение, тошнота и другие проявления, характерные длякинетоза- "морской болезни"). Сильное раздражение вестибулярного анализатора приводит к кинетозам в результате расхождения между зрительными и вестибулярными сигналами.
Афференты от полукружных каналов вместе с мозжечком и ядрами глазодвигательных нервов обеспечивают зрительный контактс окружающей средой при движении головы.
В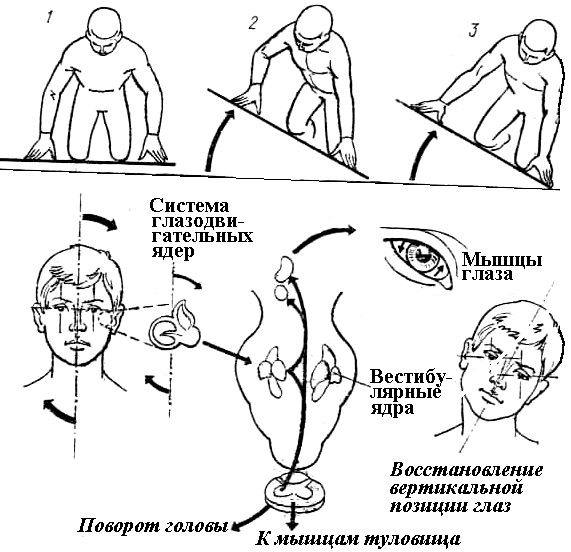 результате сопряжения активности
нейронов вестибулярного нерва (это
главным образом функция отолитового
аппарата) с двигательными ядрами ствола
и мозжечка зарождаются многие моторные
рефлексы, направленные наподдержание
позы.
результате сопряжения активности
нейронов вестибулярного нерва (это
главным образом функция отолитового
аппарата) с двигательными ядрами ствола
и мозжечка зарождаются многие моторные
рефлексы, направленные наподдержание
позы.
Рис. 6.11. Пример рефлексов, реализуемых с участием вестибулярного анализатора:1- рефлекс «сгибание-разгибание»,2- рефлекс перераспределения тонуса мышц для сохранения устойчивости,3- возвращение вертикальной оси глаз при изменении наклона головы (внизу показаны рефлекторные пути этого рефлекса).
Однако, одни лишь афферентные импульсы от вестибулярных рецепторов не в состоянии точно обеспечить предтавление о положении тела в пространстве, поскольку угол поворота головы из-за подвижности в шейном сочленении не всегда соответствует положению корпуса. Поэтому при зарождении моторных рефлексов в центрах ствола мозга, наряду с рецепцией вестибулярных нервов, используется афферентация шейных проприорецепторов, информирующих о положении головы по отношению к туловищу (подробнее об этих рефлексах сказано в гл. 7).
Проходящие через таламус импульсы от вестибулярного анализатора поступают в постцентральную извилину, где обеспечивается осознание поступившей информации, а значит и сознательная ориентация в пространстве.
Чувствительность вестибулярной системы очень велика. Нижняя граница восприятие линейных ускорений 2 см/с, а угловых 2-3о/с.
6.11. Физиология слуха
6.11.1. Рецепторы
Звук можно представить как колебательные движения упругих тел, распространяющиеся в различных средах в виде волн. Для восприятия звуковой сигнализации сформировался еще более сложный, чем вестибулярный, рецепторный орган. Формировался он рядом с вестибулярным аппаратом и поэтому в их строении есть много схожих структур. Костный и перепончатый каналы у человека образуют 2,5 витка (рис. 6.8). Слуховая сенсорная система для человека является второй после зрения по значимости и объему информации, получаемой от внешней среды.
Рецепторы слухового анализатора относятся к вторичночувствующим. Рецепторные волосковые клетки (у них редуцирован киноцилий) образуют кортиев орган, находящийся в улитке внутреннего уха, в ее средней лестнице на основной мембране, длина которой около 3,5 см. Она состоит из 20000 - 30000 волокон (рис. 6.12). Начиная от овального отверстия, длина волокон постепенно увеличивается (примерно в 12 раз), в то время как толщина их постепенно уменьшается (примерно в 100 раз).
О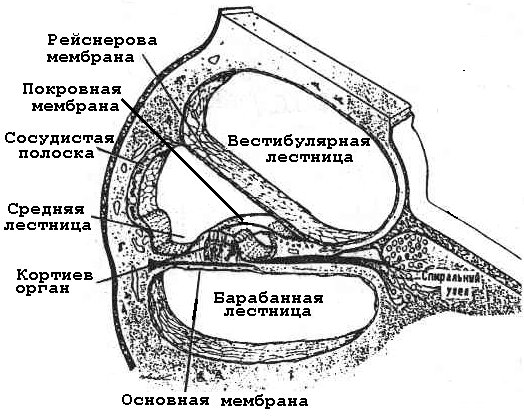 бразование
спирального (кортиева) органа завершает
текториальная мембрана, расположенная
над волосковыми клетками. На основной
мембране располагаются рецепторные
клетки двух типов:внутренниев один
ряд, анаружныев 3-4. На их мембране,
обращенной в сторону покровной мембраны,
у внутренних клеток находится 30-40
относительно коротких (4-5 мкм) волосков,
а у наружных клеток имеется 65-120 более
тонких и длинных волосков. Между
отдельными рецепторными клетками нет
функционального равенства. Об этом
свидетельствует и морфологическая
ха-рактеристика: сравнительно небольшое
количество (около 3500) внутренних клеток
дает 90% афферентов кохлеарного нерва;
в то время как от 12000 - 20000 наружных клеток
отходит лишь 10 % нейронов. Кроме того,
клетки базального и особенно среднего
витков улитки снабжены нервными
окончаниями волосковые более богато,
чем вер-хушечного витка.
бразование
спирального (кортиева) органа завершает
текториальная мембрана, расположенная
над волосковыми клетками. На основной
мембране располагаются рецепторные
клетки двух типов:внутренниев один
ряд, анаружныев 3-4. На их мембране,
обращенной в сторону покровной мембраны,
у внутренних клеток находится 30-40
относительно коротких (4-5 мкм) волосков,
а у наружных клеток имеется 65-120 более
тонких и длинных волосков. Между
отдельными рецепторными клетками нет
функционального равенства. Об этом
свидетельствует и морфологическая
ха-рактеристика: сравнительно небольшое
количество (около 3500) внутренних клеток
дает 90% афферентов кохлеарного нерва;
в то время как от 12000 - 20000 наружных клеток
отходит лишь 10 % нейронов. Кроме того,
клетки базального и особенно среднего
витков улитки снабжены нервными
окончаниями волосковые более богато,
чем вер-хушечного витка.
Рис. 6.12. Схема кортиевого органа (по Guyton).
Пространство средней лестницы заполнено эндолимфой. Над вестибулярной и под основной мембранами пространство соответствующих каналов заполненоперилимфой. Она сообщается не только с перилимфой вестибулярного трак-та, но и с субарахноидальным прос-транством мозга. Состав ее весьма близок ликвору. Эндолимфа отли-чается от перилимфы, в первую очередь тем, что в ней в 100 раз больше К+и в 10 раз меньше Nа+. То есть, по концентрации указанных ионов эти жидкости отличаются как внутриклеточная от межклеточной. Эти и другие отличия эндолимфы являются результатомактивной функции эпителия сосудистой полоски, находящейся на боковой стенке средней лестницы. Большую роль в поддержании ионного состава эндолимфы играет функция ионных насосов сосудистой полоски. Их функциональная активность сходна с эпителием почечных канальцев, и применение некоторых мочегонных препаратов может приводить к нарушению ионного состава эндолимфы и глухоте. Такой состав эндолимфы способствует повышению чувствительности рецепторного аппарата и поэтому снижение активности этих клеток приводит к ухудшению слуха.