

6.10. Чувство равновесия (физиология вестибулярного анализатора)
6.10.1. Общие представления о чувстве равновесия
В ориентации человека в пространстве помимо проприорецепторовбольшую роль играетвестибулярная сенсорнаясистема. Она информирует ЦНС о положении головы, ее движении, причем как активном, так и пассивном.
Рецепторный отдел ее - вестибулярный орган является одним из частей перепончатого лабиринта внутреннего уха (рис. 6.8).
Перепончатый лабиринт заполнен одной жидкостью, называемой эндолимфой, а погружен в другую - перилимфу.Эндолимфаимеет более высокую плотность: вязкость ее в 2-3 раза больше, чем у воды. Вестибулярный орган состоит из двух частей:статолитового аппарата, образованного маточкой и мешочком, итрех полукружных каналов. Филогенетически более юный, чем предыдущие сенсорные системы, возникший для восприятия сложного воздействия, каковым является действие сил земного притяжения, вестибулярный рецептор и устроен значительно сложнее их. В эмбриональном периоде вестибулярный аппарат закладывается рано - уже на 4-й неделе. Однако, у детей первых лет жизни возбудимость его ниже, чем у взрослых. В первые годы жизни ребенка чувствительность его повышается постепенно, а в период между 10 и 20 годами более резко.
Р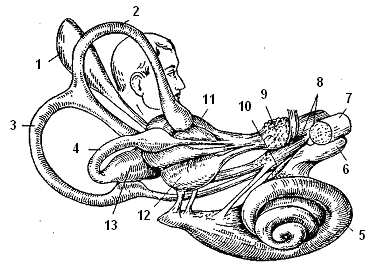 ецепторы
вестибулярной сенсорной системы
относятся квторичночувствующим
механорецепторам. В них перед
чувствительными нейронами имеются
специальные рецепторные клетки, в
которых при действии соответствующего
раздражителя возникаетвозбуждающий
рецепторный потенциал (ВРП). На
афферентный нейрон возбуждение передается
с помощью выделяющегося из этой клеткимедиатора - ацетилхолина
ецепторы
вестибулярной сенсорной системы
относятся квторичночувствующим
механорецепторам. В них перед
чувствительными нейронами имеются
специальные рецепторные клетки, в
которых при действии соответствующего
раздражителя возникаетвозбуждающий
рецепторный потенциал (ВРП). На
афферентный нейрон возбуждение передается
с помощью выделяющегося из этой клеткимедиатора - ацетилхолина
Рис. 6.8.Схема расположения вестибулярного и слухового аппарата: 1 - эндолимфатический мешочек, 2, 3, 4 - полукружные каналы, 5 - улитка, 6 - улитковый нерв, 7 - лицевой нерв, 8 - вестибулярный нерв, 9 - верхний вестибулярный узел, 10 - нижний вестибулярный узел, 11 - овальный мешочек, 12 - круглый мешочек, 13 - ампула полукружного канала.
Причем, к рецепторной клетке подходит и эфферентное волокно, которое регулирует функциональное состояние клетки, а значит, и ее чувствительность. Во время продолжительного действия раздражителя примерно две трети рецепторов вестибулярного анализатора могут адаптироваться(частота отходящих от них импульсов постепенно снижается). Хотя процессы адаптации, как и чувствительность, у данных рецепторов значительно менее выражены, чем, к примеру, у кожных, и у различных людей весьма вариабельны.
Реснитчатого типа рецепторные клетки (рис. 6.9) вестибулярного органа сгруппированы в пяти местах: втрех ампулах полукружных каналах и макулахматочки и мешочка. Каждая клетка содержит один длинный наиболее подвижный волосок, называемыйкиноцилией, и 50-60 более коротких –стероцилий,
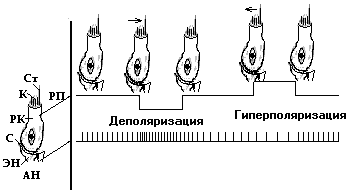
Рис. 6.9.Схема формирования электрического ответа вторичного рецептора:РП- рецепторный потенциал, ПД - потенциал действия,РК- рецептор с эффекторным (Э) и афферентным (АН)нервным окончанием, С - синапс,К- киноцилий,Ст. - стероцилии. Стрелками обозначено направление раздражителя (по Флоку).
По вестибулярному нерву от рецепторной клетки постоянно отходят ПД с частотой около 200 имп/с (фоновая импульсация).При сгибании пучка коротких ресничек в сторону киноцилия в мембране клетки повышается проницаемость для Nа+. Это приводит к деполяризации - возникновению ВРП, следствием чего в синапс, имеющийся между клеткой и афферентным волокном, выделяетсямедиатор - ацетилхолин. Взаимодействие АХ схолинорецепторомобеспечивает появление в постсинаптической мембране афферентного волокнагенераторного потенциала (ГП), который при суммации переходит в ПД. Так как и в условияхотсутствия действия раздражителя от рецепторных клеток вестибулярного анализатора постоянно отводятся ПД, топри наклоне ресничек в сторону длинного волоска частота ПДувеличивается. В отличие от этогопри наклоне ресничек в противоположную сторону частота ПДснижается. Таким путем в обоих случаях ЦНС оказывается "осведомленной" об изменении воздействия раздражителя.