

Экзамен / GISTA_EKZ_OTV
.pdfгигантских клеток костного мозга - мегакариоцитов. Ход развития мегакариоцитов из стволовой клетки крови описывается последовательностью:
СКК -> КОЕ-ГЭММ -> КОЕ-Мег -> мегакариобласт -> промегакариоцит -> мегакариоцит -> тромбоцит.
СКК, КОЕ-ГЭММ, КОЕ-Мег – это активно делящиеся клетки, морфологически схожи друг с другом, отличающиеся в основном активностью пролиферации и ответом на различные стимулирующие факторы.
Мегакариобласт – клетка диаметром 15-25 мкм, имеет ядро с инвагинациями и относительно небольшой ободок базофильной цитоплазмы. Клетка способна к делению митозом, иногда содержит два ядра. При дальнейшей дифференцировке утрачивает способность к митозу и
делится путем эндомитоза, при этом увеличиваются плоидность и размер ядра.
Промегакариоцит – клетка диаметром 30-40 мкм, содержит тетраплоидные, октоплоидные ядра
(4п, 8п), несколько пар центриолей. Объем цитоплазмы возрастает, в ней начинают накапливаться азурофильные гранулы. Клетка также способна к эндомитозу и дальнейшему увеличению полиплоидности ядер.
Мегакариоцит – дифференцированная форма. Среди них различают резервные клетки, не
образующие пластинок, и зрелые активированные клетки, образующие кровяные пластинки. при созревании из мегакариобласта становится крупнее, достигая 50-100 мкм в диаметре; его
ядро и цитоплазма претерпевают выраженные изменения.
Дифференцировка ядра включает активную репликацию ДНК без митоза. Полиплоидные клетки в дальнейшем претерпевают эндомитоз с образованием многочисленных связанных перемычками долей ядра, в котором общее содержание ДНК соответствует 4-128n (наиболее часто - 16 или 32n).
Хроматин постепенно конденсируется, ядрышко, как правило, не выявляется. Размеры клетки обычно соответствуют степени ее полиплоидизации.
Дифференцировка цитоплазмы мегакариоцитов начинается только по завершении репликации ДНК. Наиболее заметными ее проявлениями служат:
●Разделение цитоплазмы на три зоны:
○Околоядерная зона содержит элементы грЭПС, хорошо развитый комплекс Гольджи, митохондрии и центриоли.
○Промежуточная зона - наибольшая по ширине, содержит гранулы и систему мембранных демаркационных каналов – каналов, разрезающих цитоплазму мегакариоцитов на территории размером 2-4 мкм, соответствующие границам будущих тромбоцитов и содержащие гранулы.
○Краевая (периферическая) зона свободна от большинства органелл и гранул, в ней в значительном количестве сосредоточены элементы цитоскелета .
●Образование и накопление гранул, характерных для тромбоцитов и содержащих типичные для них белки
●Образование протромбоцитов – узких длинных лентовидных отростков мегакариоцитов,
которые через поры эндотелия синусов красного костного мозга проникают в их просвет, где распадаются на отдельные тромбоциты.
41
18. Моноцитопоэз. Основные стадии развития и дифференцировки моноцитов.
Моноцитопоэз - процесс развития моноцитов - происходит в красном костном мозге и описывается последовательностью:
СКК -> КОЕ-ГЭММ -> КОЕ-ГМ -> КОЕ-М -> монобласт -> промоноцит -> моноцит
СКК, КОЕ-ГЭММ, КОЕ-ГМ, КОЕ-М – это активно делящиеся клетки, морфологически схожи друг с другом, отличающиеся в основном активностью пролиферации и ответом на различные стимулирующие факторы.
Промоноцит - сравнительно крупная клетка (диаметром 12-18 мкм) с большим светлым слегка вогнутым ядром, в котором располагаются 1-2 ядрышка. Базофильная цитоплазма содержит
умеренно развитую грЭПС, полисомы, митохондрии, центриоли и крупный комплекс Гольджи, от которого отделяются незрелые азурофильные гранулы.
Промоноциты делятся и постепенно дифференцируются в моноциты.
Процесс преобразования монобластов в моноциты включает:
●Дальнейшее увеличение размеров клетки преимущественно за счет нарастания объема цитоплазмы
●Снижение базофилии цитоплазмы
●Накопление в ней (лизосом)
●Изменение формы ядра, которое становится бобовидным
+
Выделяясь в синусы ККМ, они попадают в кровь, в которой циркулируют от 8 часов до 3-4 суток,
а далее через стенку сосудов мигрируют в ткани. Лишь около 5% моноцитов, имеющихся в организме, циркулирует в крови, остальные находятся во внесосудистое пуле. В тканях они превращаются в различные виды макрофагов (вместе с которыми образуют единую моноцитарномакрофагальную систему), а также в дендритные антиген-представляющие клетки
19.Т-лимфоциты: субпопуляции. Характеристика рецепторов. Антигеннезависимая и антигензависимая пролиферация и дифференцировка.
Т-лимфоциты — самая многочисленная популяция лимфоцитов, составляющая 70—90 % лимфоцитов крови. Эти клетки имеют на своей плазмолемме маркеры , позволяющие идентифицировать их, а также ТКР (Т-клеточные рецепторы) – специфические антигенраспознающие рецепторы.
Субпопуляции Т-лимфоцитов:
●Т-хелперы – стимулируют дифференцировку В-лимфоцитов, образование из них плазмоцитов и продукцию иммуноглобулинов. CD3 + CD4 + ТКР
●Т-супрессоры – способны ингибировать действие Т-хелперов, В-лимфоцитов, плазмоцитов и регулировать силу иммунного ответа. CD3 + CD8 + ТКР
●Т-киллеры – участвуют в реакциях клеточного иммунитета, обеспечивая разрушение (лизис) чужеродных клеток и собственных измененных клеток (например, опухолевых клеток). CD3 + CD8 + ТКР
42
●Т-памяти – долгоживущие клетки, которые активируются при повторном попадании чужеродного объекта в организм. CD45RO+ и ТКР. Клетка памяти на своей плазмолемме имеет много ТКР, потому что должна активироваться даже при маленьких концентрациях антигена.
Антигеннезависимая пролиферация.
Протимоцит (про-Т-лимфоцит) образуется в красном костном мозге из КОЕ-Л и соответствует стадии, предшествующей реаранжировке генома.
Пре-Т-клетки мигрируют из костного мозга через кровь в центральный орган иммунной системы – вилочковую железу. Мигрирующие в вилочковую железу пре-Т-клетки приобретают способность реагировать на стимулы микроокружения.
Незрелые Т-лимфоциты и зрелые Т-лимфоциты - дозревают и дальше начинается антигензависимая пролиферация и последовательные стадии, идущие за претимоцитами. Эти клетки претерпевают реаранжировку генома с формированием разнообразных специфических антигенраспознающих Т Клеточные Рецепторы, которые экспрессируются на их поверхности. На плазмолемме появляется ряд маркеров, типичных для зрелых Т-лимфоцитов и необходимых для их взаимодействия с другими клетками.
Антигензависимая пролиферация:
Покидая тимус, наивные (зрелые) Т-лимфоциты с током крови мигрируют в Т-зависимые зоны периферических органов кроветворения и иммуногенеза. В этих органах они встречаются с
антигенами, которые им представляют АнтПредстКлетк после процессинга, и взаимодействуют с Т-хелперами.
Взаимодействуя с антигеном, Т-лимфоциты активируются, подвергаются бласт-трансформации –
превращаются в Т-иммунобласты. Последние пролиферируют и дифференцируются, формируя крупные клоны эффекторных и регуляторных клеток. Часть Т-лимфоцитов превращается в долгоживущие Т-клетки памяти.
20 Механизм активации Т-лимфоцитов в условиях иммунологической реакции клеточного типа. Этапы взаимодействия Т-киллера с клеткой-мишенью.
Иммунологическая реакция клеточного типа – специфический защитный механизм,
осуществляющийся путем непосредственного контактного взаимодействия клеток-эффекторов иммунной системы с клетками-мишенями, несущими чужеродные или измененные собственные антигены. Наблюдается при трансплантации органов и тканей, злокачественном опухолевом росте, инфицировании вирусами.
В клеточном иммунитете участвует Т-киллер, реагирующий с антигеном в комплексе с гликопротеинами МНС I класса в плазматической мембране клетки-мишени. Т-киллер активируется, если узнает с помощью своих рецепторов фрагменты чужеродных белков,
связанные с молекулами МНС I класса на поверхности зараженной клетки, или если на
поверхности клетки-мишени отсутствует МНС-I класса.
Взаимодействие Т-киллера с клеткой-мишенью проходит в несколько этапов:
43

1.Контакт Т-киллера с клеткой-мишенью
2.Увеличение в Т-лимф концентрации ионов Са 2+
3.Экзоцитоз гранул перфорина в межклеточное пространство
4.Встраивание мономеров перфорина в плазмалемму клетки-мишени
5.Формирование трансмембранных пор за счет полимеризации белка перфорина
6.Нарушение осмотического равновесия в клетке-мишени, набухание и ее гибель.
21 В-лимфоциты (субпопуляции). Характеристика рецепторов, антигеннезависимая и антигензависимая пролиферация и дифференцировка.
В-лимфоциты отличаются от других типов клеток способностью синтезировать иммуноглобулины. Зрелые В-лимфоциты экспрессируют Ig на клеточной мембране. Такие мембранные иммуноглобулины (mlg) функционируют как антигенспецифические рецепторы.
Пре-В-клетки синтезируют внутриклеточный цитоплазматический IgM, но не имеют поверхностных иммуноглобулиновых рецепторов . Зрелые В-лимфоциты несут на своей поверхности иммуноглобулиновые рецепторы различных классов — IgM, IgG, IgА.
Именно зрелые В-лимфоциты делятся на 2 субпопуляции, которые синтезируют определенный класс иммуноглобулинов.
Антигеннезависимая пролиферация и дифференцировка В-лимфоцитов осуществляется в ККМ.
Про-В-лимфоцит В цитоплазме выявляется IgМ, но он отсутствует на его плазмолемме; для незрелого В-лимфоцита характерна экспрессия на плазмолемме.
Развитие В-лимфоцитов сопровождается утратой одних клеточных маркеров и приобретением других, в частности, функционально важных в процессах адгезии, активации и рецепции цитокинов маркеров CD19, СD20, CD21, СD22, СD23 и СD40, рецепторов к комплементу, Fсфрагменту иммуноглобулинов. В зрелом В-лимфоците он экспрессируется совместно с IgD.
44
Антигензависимая пролиферация В-лимфоцитов
+
Покидая красный костный мозг, зрелые В-лимфоциты, на поверхности которых экспрессируются
IgM и IgD циркулируют в крови и попадают в периферические органы кроветворения и иммуногенеза. В этих органах они взаимодействуют с антигеном, соответствующим по специфичности их рецепторам, а также с Т-хелперами и активируются, подвергаясь
бласттрансформации и превращаясь в течение 1-2 сут. в В-иммунобласты. Последние спустя
3-4 сут. дают начало плазмобластам (далее дифференцирующимся в плазматические клетки) и В-
клеткам памяти.
22.Антигены. Растворимые и нерастворимые. Гуморальный иммунитет. Фазы иммунного ответа.
Антигены – вещества, несущие генетически чужеродную информацию. Антигены бывают 2 типов:
●Растворимые. Например, белки, полисахариды, нуклеопротеиды
●Нерастворимые. Например, опухолевые клетки, клетки зараженные вирусом, трансплантируемые ткани или органы.
Гуморальный иммунитет – специфический защитный механизм, который обеспечивается путем выработки антител, переносимых с кровью и тканевыми жидкостями. Разыгрывается на растворимый антиген. За реакцию этого типа отвечают АнтПредстКлетк, т-хелперы и В-
лимфоциты (которые дифференцируются в плазматические клетки).
Фазы иммунного ответа:
1.Поглощение антигена. Макрофаг (или другая АПК) движется в сторону АГ, реагируя на положительные раздражители, здесь наблюдается положительный хемотаксис
2.Процессинг( обработка) антигена системой гидролитических ферментов макрофагов до низкомолекулярных фрагментов
3.Представление антигена. Образовавшиеся комплексы белок МНС II + антигенный пептид (включает 12-25 аминокислотных остатков) экспрессируются на мембране и представляются Т-хелперам (1-й сигнал). Одновременно макрофаг активируется и выделяет фактор (ИЛ-1), который также стимулирует Т-хелперы (2-й сигнал).
4.Индуктивная фаза. Т-хелперы, получив два сигнала от макрофагов, выделяют ИЛ-2, который стимулирует пролиферацию Т-лимфоцитов и В-лимфоцитов.
5.Эффекторная стадия. Активированные Т-клетки экспрессируют рецептор к ИЛ-2 и CD40L. Последний взаимодействует с CD40 рецептором на В-клетках и возникает взаимная стимуляция этих клеток. В-лимфоциты превращаются в плазматические клетки
(запускается процесс бласт-трансформации), которые синтезируют антитела, специфичность которых увеличивается у потомков делящихся клеток.
Антитела осуществляют
Антитоксический эффект ― нейтрализацию токсинов бактерий и вирусы путем их связывания
●Нейтрализацию бактерий и вирусов путем взаимодействия с рецепторами клетки, связывающими бактерии или вирусы, препятствуя тем самым их адгезии и проникновению в клетку
45
●Опсонизацию ― повышение фагоцитарной активности лейкоцитов (т.е. фагоцитирующие лейкоциты начинают фагоцитировать связанные антигены)
23. Классификация иммунокомпетентных клеток. Клеточный иммунитет. Фазы иммунного ответа
Функциональная классификация иммунокомпетентных клеток:
1.Антигенпредставляющие клетки(АПК) – захватывают антигены , перерабатывают их и представляют их другим иммунокомпетентным клеткам. (Макрофаги, дендритные клетки и т.д.)
2.Эффекторные клетки, непосредственно осуществляющие реакции иммунитета (плазмоциты, Т-киллеры)
3.Регуляторные клетки, обеспечивающие активацию или угнетение отдельных звеньев иммунных реакций (Т-хелперы, Т-супрессоры)
4.Клетки-памяти, хранящие информацию о взаимодействии конкретным антигеном и тем самым способствующие более активному развитию иммунного ответа при повторном его воздействии (Т- и В-памяти)
Морфологическая классификация иммунокомпетентных клеток:
1.Дендритные антигенпредставляющие клетки (АПК), которые захватывают антигены и представляют их лимфоцитам в переработанном виде, обусловливая тем самым "запуск" иммунных реакций
2.Лимфоциты - основные клетки, обеспечивающие развитие и течение иммунных реакций
- путем непосредственного участия в них (эффекторные клетки) или регуляторных воздействий на другие клетки. В-лимфоциты способны выполнять роль АнтПредстКлеткой .
Лимфоциты образуют несколько функционально специализированных групп (субпопуляций), постоянно мигрируют (рециркулируют) в организме и взаимодействуют как друг с другом, так и с клетками других типов посредством адгезивных контактов и цитокинов.
3.Макрофаги, которые, наряду с участием в реакциях неспецифической защиты, могут выполнять функции как АПК, так и эффекторных клеток иммунных реакций.
Клеточный иммунитет – специфический защитный механизм, осуществляющийся путем непосредственного контактного взаимодействия клеток-эффекторов иммунной системы с клетками-мишенями, несущими чужеродные или измененные собственные антигены. Данный механизм разыгрывается на нерастворимый антиген.Клеточный иммунитет направлен
преимущественно против микроорганизмов, которые выживают в фагоцитах и против микроорганизмов, поражающие другие клетки. Система клеточного иммунитета особенно эффективна против клеток, инфицированных вирусами, и принимает участие в защите от грибов, простейших, внутриклеточных бактерий и против клеток опухолей. Также система клеточного
иммунитета играет важную роль в отторжении тканей.
Клеточный иммунный ответ включает в себя следующие фазы:
Фаза распознавания нерастворимого антигена
1.Эндоцитоз антигена АПК
2.Процессинг (обработка ) антигена, т.е. его частичное расщепление с сохранением высокоиммуных антигенных детерминант, имеющих вид линейных пептидных цепочек, длиной 12-15 аминокислотных остатков.
46
3.Формирование комплекса эпитоп с молекулой главного комплекса гистосовместимости (МНС) и появление его на поверхности АПК
4.Презентация комплекса эпитоп +МНС 1 класса на поверхности АПК
Фаза активации Т-лимфоцитов
1. Началом этой фазы являются два сигнала, которые АПК подаёт Т-хелперу:
●Комплекс эпитопа антигена с молекулой МНС 1 класса
●Синтез интерлейкинов
Т-хелперы стимулируют бласттрансформацию Т-киллеров
+
Фаза взаимодействия Т-киллера с клеткой-мишенью
1.Контакт Т-киллера с клеткой-мишенью
2.Увеличение в Т-лимф концентрации ионов Са 2+
3.Экзоцитоз гранул перфорина в межклеточное пространство
4.Встраивание мономеров перфорина в плазмолемму клетки-мишени
5.Формирование трансмембранных пор за счет полимеризации белка перфорина
6.Нарушение осмотического равновесия в клетке-мишени, набухание и ее гибель
24.Рыхлая волокнистая соединительная ткань. Клеточный состав, источники развития и обновления. Механизмы связи клеток с компонентами межклеточного вещества
Рыхлая волокнистая соединительная ткань – вид соединительной ткани, имеющий большое количество гелеобразного межклеточного вещества с небольшим количеством разнонаправленных волокон. Обнаруживается во всех органах, так как она сопровождает кровеносные и лимфатические сосуды, нервы и образует межтканевые прослойки многих органов.
Источники развития и обновления.
РВСТ, как и все соединительные ткани образуется из мезенхимы, причём сам процесс образования в разных органах проходит неодинаково и зависит от неодинаковой физиологической значимости на разных этапах эмбриогенеза. Развитие РВСТ связано с высокой скоростью размножения клеток,
волокнообразования, постоянной перестройкой ткани (резорбцией и новообразованием).
Обновление происходит медленнее и направлено на поддержание тканевого гомеостаза, пролиферацию малодифференцированных клеток и замену или отмирающих клеток.
Основными клетками РВСТ являются фибробласты (все его диффероны), макрофаги, тучные клетки, адвентициальные клетки, плазматические клетки, перициты, жировые клетки, а также лейкоциты, мигрирующие из крови.
Фибробласты будут в вопросе 25!!
Макрофаги (в РВСТ называются гистиоциты) – это клетки, выполняющие защитную функцию, прежде всего посредством фагоцитоза крупных частиц. Макрофаги образуются из моноцитов
после выхода последних из кровеносного русла.
Тучные клетки регулируют местный тканевой гомеостаз посредством выработки веществ,
способных изменять проницаемость гемокапилляров и степень гидратации межклеточного вещества (гистамин, гепарин, серотонин), а также принимают участие в иммунных реакциях.
47
Адвентициальные клетки – локализуются в адвентиции сосудов. Адвентициальные клетки являются камбиальными полипотентными предшественниками фибробластов, остеобластов и адипоцитов.
Плазматические клетки являются эффекторными клетками гуморального иммунитета. Они
образуются из В-лимфоцитов при воздействии на них антигенов. Функции плазмоцитов – синтез и
выделение иммуноглобулинов.
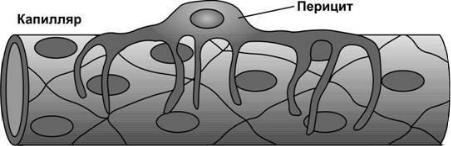
Перициты – клетки отросчатой формы, которые прилегают к стенке капилляра лишь с одной стороны и охватывают его в виде корзинки. Перициты контролируют пролиферацию эндотелия,
синтезируют компоненты базальной мембраны, а также способны дифференцируется в гладкие миоциты и фибробласты, осуществляя таким образом репаративную функцию.
Жировые клетки (адипоциты) – это клетки, способные в больших количествах накапливать резервный жир. Развиваются из адвентициальных клеток.
Механизмы связи клеток с компонентами межклеточного вещества.
Белок синдекан вместе с мембранным рецептором CD44 с внутренней стороны связываются с цитоскелетом, а с наружной стороны связывают поверхность клетки с фибронектином.
Аналогичную функцию выполняют интегрины – адгезивные гликопротеины, которые через белки таллин и винкулин связывают актиновые микрофиламенты цитоскелета с коллагеновыми волокнами
– непосредственно или опосредованно – через молекулы фибронектина.
+
Белок перлекан, связывается с фибронектином и опосредует прикрепление фибробласта к компонентам межклеточного вещества.
25 .Дифферон фибробластов. Световая и электронная микроскопия типов клеток. Функции фибробластов.
Фибробластический дифферон включает в себя стволовые клетки, полустволовые клеткипредшественники, малодифференцированные (юные), дифференцированные (зрелые) фибробласты, фиброциты, а также фиброкласты и миофибробласты. Источником развития фибробластов являются адвентициальные клетки. В рыхлой волокнистой соединительной ткани фибробласты располагаются свободно в основном веществе, не образуя межклеточных контактов друг с другом.
1.Малодифференцированный (юный) фибробласт - базофильная клетка (20-25 мкм), с
небольшим числом отростков. Для нее характерно крупное круглое или овальное ядро с 1-2 ядрышками, умеренно развитый синтетический аппарат. Она сохраняет способность к
48
пролиферации, но они уже начинает осуществлять синтез типичных компонентов
межклеточного вещества соединительной ткани - коллагена и гликозаминогликанов.
Способность юных фибробластов к направленной миграции определяет их важную роль в репаративных процессах, в частности, в заживлении ран. Миграция осуществляется благодаря наличию в их цитоплазме сократимых микрофиламентов, на которые опосредованно передаются сигналы с многочисленных рецепторов плазмолеммы, воспринимающих молекулы хемотаксических веществ.
2.Дифференцированные (зрелые) фибробласты являются центральным звеном фибробластического дифферона. Крупная (более 40-50мкм) отростчатая клетка со светлым ядром, содержащим мелкодисперсный хроматин и 1-2 ядрышка. Цитоплазма делится на
эндоплазму (содержит большую часть органелл мощно развитого синтетического аппарат для синтеза межклеточного вещества) и эктоплазму (заполнена преимущественно элементами цитоскелета). Фибробласт обладает подвижностью, способностью изменять
свою форму и обратимо прикрепляться к другим клеткам и компонентам межклеточного вещества.
3.Фиброциты – конечная форма развития фибробласта - узкая веретенообразная, неспособная к пролиферации клетка с длинными тонкими отростками, которые часто имеют уплощенную крыловидную форму. Ядро - сравнительно плотное (с преобладанием гетерохроматина),
занимает большую часть клетки. Цитоплазма содержит слабо развитый синтетический аппарат, т.е. синтез компонентов межклеточного вещества осуществляется ими очень слабо.
4.Миофибробласты – особые клетки, которые по своему строению и функции занимают
промежуточное положение между типичными фибробластами и клетками гладкой мышечной ткани - гладкими миоцитами. Они активно участвуют в процессах заживления
раны, стягивая края и образуя коллаген.
5.Фиброкласты – клетки дифферона фиброцитов, специализированные на функции
разрушения межклеточного вещества соединительной ткани, которая резко преобладает над их синтетической и секреторной активностью. В их цитоплазме выявляются многочисленные вакуоли, содержащие литические ферменты и коллагеновые фибриллы на различных стадиях лизиса. Эти клетки обеспечивают перестройку и инволюцию соединительной ткани
Функции фибробластов:
1.Образования межклеточного матрикса (в основном этим занимаются фибробласты)
2.Образуют коллагеновые и эластиновые волокна
3.Гликозаминогликаны и протеогликаны, формирующие основное вещество
4.Поддержания нормального гомеостаза межклеточного вещества (в основном этим занимаются фиброциты)
5.Секретируют цитокины, регулирующие кроветворение: колониестимулирующие факторы макрофагов и гранулоцитов
6.Участвуют в заживлении ран
26.Макрофаги (гистиоциты) и плазматические клетки (СМ и ЭМ), участие в иммунных реакциях.
Макрофаги (гистиоциты) - вторые по численности (после фибробластов) клетки рыхлой волокнистой соединительной ткани. Они принадлежат к линии потомков стволовой клетки крови и непосредственно образуются из моноцитов после их миграции в соединительную ткань из просвета кровеносных сосудов. В соединительной ткани макрофаги располагаются поодиночке или
49
группами. Эти клетки очень многочисленны в собственной пластинке слизистых оболочек, а
также в серозных оболочках. Они могут пребывать в одном из двух взаимообратных состояний:
1.Покоящихся клеток, обладающих низкой функциональной активностью
2.Блуждающих клеток с высокой функциональной активностью
Покоящиеся гистиоциты трудно идентифицировать на светооптическом уровне. Они имеют вид
мелких уплощенных клеток удлиненной или отросчатой формы с четкими контурами, прикрепленных к коллагеновым волокнам. Эти клетки характеризуются небольшим земным ядром и плотной цитоплазмой со слабо развитыми органеллами. Могут превращаться в
блуждающие под действием микроорганизмов или их продуктов, а также ряда цитокинов
Блуждающие (активные) гистиоциты обладают высокой подвижностью, изменчивой (отросчатой, реже округлой) формой с неровными, но обычно четко выявленными краями. Их ядро светлее, чем в покоящихся клетках, но темнее, чем в фибробластах; в нем может выявляться ядрышко. Цитоплазма содержит многочисленные лизосомы и развитые элементы цитоскелета,
которые концентрируются в области псевдоподий; другие органеллы развиты умеренно.
+
В иммунных реакциях макрофаги выполняют роль АПК, т. е. участвуют в иммунных реакциях посредством захвата, процессинга антигена, предоставления комплекса [эпитоп + МНС класса 2] Т-
хелперам и их активация. Также они синтезируют цитокины, стимулирующие пролиферацию
лимфоцитов и вовлечение их в иммунную реакцию.
Плазматические клетки (плазмоциты) и их предшественники В лимфоциты, находящиеся на различных этапах преобразования в плазмоциты - в небольших количествах постоянно содержатся в различных участках рыхлой волокнистой соединительной ткани. Это неподвижные или очень слабо подвижные, короткоживущие. Плазматические клетки имеют сравнительно мелкие размеры (диаметр 9-20 мкм, в среднем - 14 мкм), круглую или овальную
форму. Ядро - округлое, расположено эксцентрично, содержит крупные глыбки гетерохроматина, которые располагаются в виде радиальных тяжей ("спиц колеса"). Ядрышко крупное, лежит в центре ядра или эксцентрично. В отличие от других клеток, интенсивно вырабатывающих белок, продукты синтетической деятельности плазматических клеток (иммуноглобулины) в норме не накапливаются в цитоплазме в секреторных гранулах, а по мере образования транспортируется мелкими пузырьками к плазмолемме, где непрерывно выделяется механизмом экзоцитоза.
Функция плазматических клеток заключается в обеспечении гуморального иммунитета путем выработки антител.
27.Биосинтез коллагена. Уровни структурной организации коллагеновых и эластических волокон. Типы коллагена, зоны локализации в организме.
Молекулы коллагена построены из триплетов – трех пептидных цепочек (а(альфа)-цепочки) –
проколлагена, свивающихся ещё в клетке в единую спираль.
Этот процесс проходит в клетке в несколько этапов:
1.Синтез а-цепи проколлагена
2.Гидроксилирование (присоединение –ОН груп) части остатков пролина и лизина
50