

Экзамен / GISTA_EKZ_OTV
.pdf1-21, 22-77, 78-174
ЦИТОЛОГИЯ С ЭМБРИОЛОГИЕЙ
1.Структурно-химический состав и молекулярная организация плазмолеммы.
Плазмолемма – это поверхностная периферическая структура клетки, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой.
Основу плазмолеммы составляет липопротеиновый комплекс . Она имеет толщину 8-10 нм. Основными химическими компонентами клеточных мембран, в частности плазмолеммы, являются белки, липиды и углеводы.
Снаружи от плазмолеммы располагается надмембранный слой – гликокаликс – ассоциированный с плазмолеммой гликопротеиновый комплекс, в состав которого входят углеводы. Эти углеводы имеют различные функции, а также придают мембране отрицательный заряд. Толщина этого слоя варьируется в пределах 5-100 нм
Изнутри располагается кортикальный (подмембранный) слой, содержащий элементы цитоскелета. Толщина этого слоя варьируется в пределах 20-40 нм
К липидам плазматической мембраны относится большое количество гидрофобных веществ.
Основные представители – фосфолипиды, холестерин, сфингомиелин (в основном в цнс). Фосфолипиды располагаются в 2 слоя и имеют 2 части – гидрофильные головки, обращенные кнаружи в обе стороны и гидрофобные хвосты, обращенные внутрь. Основная функция фосфолипидов , это формирование основного каркаса биологической мембраны .
Мембранные белки также состоят из 2 частей – участков, богатых полярными аминокислотами
и обращенных кнаружи, и гидрофобных участков, находящихся в гидрофобной части мембраны. Выделяют белки:
1.Интегральные - Белки-каналообразующие и белки ферменты
2.Полуинтегральные -Белки-переносчики
3.Примембранные -. Рецепторные белки
4.Структурные белки
Углеводы мембран входят в их состав не в свободном состоянии, они связаны с молекулами липидов или белков. Такие вещества называются соответственно гликолипидами и гликопротеидами.
̆ ̆
2.Надмембранный (гликокаликс) и подмембранный (кортикальный) компоненты плазмолеммы. Особенности строения и функции. Мембранные рецепторы
Надмембранный компонент (Гликокаликс)
представляет собой ассоциированный с плазмолеммой комплекс, в состав которого
входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы. Углеводные участки
гликолипидов и гликопротеинов придают поверхности клетки отрицательный

заряд.Толщина 5-100 нм
Эти углеводные участки играют роль рецепторов (веществ, чувствительных к отдельным веществам), обеспечивающих распознавание клеткой - соседних клеток и межклеточного вещества , а также адгезивные взаимодействия с ними. ( сцепление поверхностей разнородных твёрдых и/или жидких тел).
Всостав гликокаликса некоторые авторы включают, помимо углеводных компонентов, периферические мембранные белки и полуинтегральные белки, функциональные участки которых находятся в надмембранной зоне (например, иммуноглобулины).
Вгликокаликсе находятся рецепторы гистосовместимости, некоторые ферменты, рецепторы гормонов. Такие чувствительные к отдельным веществам участки могут быть разбросаны по всей поверхности клетки или собраны в небольшие зоны.
Гликокаликс также участвует в пристеночном пищеварении ( зона адсорбции ферментов )
Выделяют рецепторы:
●Ионотропные, которые при взаимодействии с субстратом образуют специфический ионный канал. Эти рецепторы представляют собой сложный белок.
●Метаботропные – представлены гликопротеинами
●Каталитические – включают внеклеточную часть (собственно рецептор) и
цитоплазматическую часть, которая функционирует как протеинкиназа.
●Связанные с G-белками - трансмембранные белки, ассоциированные с ионным каналом или ферментом.
Состоят из:
–рецептора, взаимодействующего с сигнальной молекулой (первый посредник),
–G-белка, который передает сигнал на связанный с мембраной фермент
(аденилатциклазу) или ионный канал, вследствие чего активируется второй внутриклеточный посредник - чаще всего цАМФ или Са2+.
Подмембранный компонент (Кортикальный) образован специализированной периферической частью цитоплазмы, прилежащей к плазмолемме и содержащей элементы цитоскелета,
преимущественно актиновые микрофиламенты. Более глубоко располагаются промежуточные филаменты и микротрубочки. Благодаря сокращению сети микрофиламентов, связанных с белками плазмолеммы, происходят изменения формы клетки и ее отдельных участков, формирование псевдоподий, выростов, перемещение клетки в пространстве. Также благодаря этому комплексу происходит перемещение белковых частиц, входящих в состав плазмолеммы с одного места на другое. Толщина 20-40 нм
3.Молекулярная организация плазмолеммы. Пассивный и активный транспорт. Эндоцитоз и его разновидности.
Молекулярная организация плазмолеммы
Молекулярное строение описывается жидкостно-мозаичной моделью, согласно которой она состоит из липидного (фосфолипидного) бислоя, в который погружены и с которым связаны молекулы белков.
Пассивный транспорт – транспорт низкомолекулярных соединений по градиенту концентрации без затраты энергии. Это происходит за счёт каналов без АТФазной активности, которые или
открыты постоянно, или открываются под воздействием различных стимулов (потенциалзависимые, лиганд-зависимые). Также пассивный транспорт осуществляется путём растворения гидрофобных (или липофильных) веществ в плазмолемме.
2
Активный транспорт – перемещение веществ через мембрану против градиента концентрации с затратой энергии. Осуществляется за счёт специальных белков переносчиков.
Выделяют:
●Первично-активный транспорт, осуществляемый за счёт использования энергии макроэргической связи АТФ. Например, Na-K насос.
1)Вторично-активный транспорт, осуществляемый за счёт ранее созданного с помощью первично-активного транспорта градиента концентрации. Например, перенос аминокислот внутрь клетки за счёт котранспорта с ионом натрия.
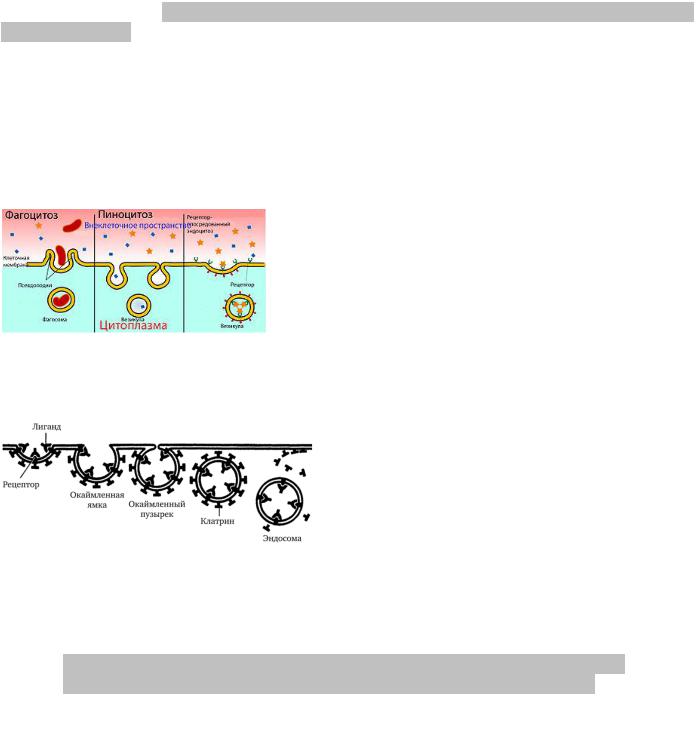
Макромолекулы, их агрегаты и крупные частицы попадают в клетку путём эндоцитоза.
Эндоцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Эти впячивания могут иметь вид еще не замкнутых округлых пузырьков
или представлять собой глубокие инвагинации, впячивания внутрь клетки.
Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
Часто для экономии (образования меньшего количества пузырьков) в ходе процесса эндоцитоза рецепторы
определенных макромолекул перемещаются латерально по плазмолемме и накапливаются в окаймленной ямке. Благодаря белку клатрину, который образует сетевидную структуру в области
этой ямки, вытесняются все остальные рецепторы из этой зоны. Через 1 минуту окаймленная
ямка превращается в окаймленный пузырёк и теряет клатриновую оболочку, после чего происходит слияние пузырька с лизосомой и начинается процессинг лигандов, а рецепторы вторично используются в плазмолемме.
Эндоцитоз условно разделяют на:
2)Пиноцитоз – захват и поглощение клеткой жидкости и/или растворимых веществ.
3)Фагоцитоз – захват и поглощение клеткой плотных, обычно крупных частиц; обычно сопровождается образованием выпячиваний цитоплазмы - псевдоподий, охватывающих объект фагоцитоза и смыкающихся над ним.
̆
4.Специализированные структуры плазмолеммы: микроворсинки, реснички, базальный лабиринт.
Дифференцированность плазмалеммы наиболее выражена у поляризованных клеток, в частности клеток эпителиев. Для таких клеток характерно наличие двух полюсов (апикального (граничит с внешней средой) и базального). В соответствии с этим в плазматической мембране выделяют
апикальную, латеральную (боковую) и базальную части.
Специализация апикальной части плазмалеммы (жгутики, реснички и микроворсинки). 1 — актиновые микрофиламенты; 2 — верхушечный элемент для прикрепления актиновых микрофиламентов; 3—5 — специальные белки
Микроворсинки – пальцевидные выросты цитоплазмы клетки диаметром 0.1 мкм и длиной 1 мкм (СМ), основу которых составляют актиновые микрофиламенты, располагающихся в
3

кортикальном слое цитоплазмы. Каркас каждой микроворсинки образован пучком, содержащим около 40 микрофиламентов, лежащих вдоль ее длинной оси. В апикальной части микроворсинки этот пучок закреплен в аморфном веществе. Его жесткость обусловлена поперечными сшивками из белков фимбрина и виллина, изнутри пучок прикреплен к плазмолемме микроворсинки особыми белковыми мостиками. У основания микроворсинки микрофиламенты пучка вплетается в терминальную сеть, среди элементов которой имеются миозиновые филаменты. (ЭМ) Микроворсинки обеспечивают многократное увеличение площади поверхности клетки, на которой происходит расщепление и всасывание веществ.
Имеются у клеток кишечного эпителия и эпителия почечных канальцев.
Функции — мембранное пищеварение и всасывание.
Реснички — короткие и многочисленные; встречаются у одноклеточных (инфузории) и некоторых клеток многоклеточных организмов (клетки эпителия трахеи)
а — схема строения: б — пучки ресничек на внутренней поверхности эпителия трахеи
Реснички – органеллы специального значения,
участвующие в процессах движения, - представляют собой выросты цитоплазмы, основу которых составляет аксонема (каркас из микротрубочек). Длина ресничек равна 2-10 мкм (ЭМ). Аксонема образована 9
периферическими парами микротрубочек и одной центрально расположенной парой; такое строение описывается формулой (9 х 2) + 2. В парах микротрубочки связаны белком динеином, который
обладает АТФазной активностью (за счёт него происходит движение реснички). В основании аксонемы лежит базальное тельце, которое является матрицей для построения аксонемы. Состоит оно из девяти триплетов микротрубочек на периферии, в центре микротрубочек нет (9х3+0) (ЭМ).
Функции:
● для подвижных клеток – перемещение, например, для сперматозоида ●для неподвижных клеток – перемещение жидкости или частиц, например,
ресничные клетки ресничного эпителия (дыхательного) перемещают частицы пыли.
Базальный лабиринт –углубление плазмолеммы базального полюса клетки с лежащими между ними митохондриями. Эти митохондрии вырабатывают энергию, которая потребляется ионными насосами в их плазмолемме. Функция эпителиоцитов, обладающих базальной исчерченностью, связана с изменением ионного состава жидкости (мочи, слюны) в просвете указанных канальцев и протоков. Форму базального лабиринта поддерживают структуры цитоскелета: промежуточные филаменты и тонкие микрофиламенты Функции — транспорт воды и различных веществ в клетку и из клетки.
5.Рибосомы. Полисомы. Синтез белка на свободных полисомах.
4

Рибосо́ма — немембранный органоид живой клетки сферической или слегка эллипсоидной формы,
Диаметр 15—20нм (прокариоты), 25—30нм (эукариоты).
Состоит из большой и малой субъединиц.
Служат для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК, или мРНК. Этот процесс называется трансляцией.
В эукариотических клетках рибосомы располагаются на мембранах эндоплазматической сети, хотя могут быть локализованы
и в неприкрепленной форме в цитоплазме.
Нередко с одной молекулой мРНК ассоциировано несколько рибосом, такая структура называется
полирибосомой (полисомой).
Синтез рибосом у эукариот происходит специальной внутриядерной структуре — ядрышке
Классификация
Клеточные рибосомы. В клетках животных различают два типа рибосом:
●цитоплазматические рибосомы, которые содержатся в цитозоле и значительно преобладают по числу, и размеру. Подразделяются на 2 подтипа:
a)свободные рибосомы — находятся в гиалоплазме, не будучи связанными с мембранами ЭПС
b)мембраносвязанные рибосомы — фиксированы на наружной (обращенной к гиалоплазме) поверхности ЭПС. В обоих случаях находятся, в составе полисом.
●митохондриальные рибосомы — содержатся в митохондриях, несколько отличаясь своим составом (более мелкие).
Строение.
В собранном виде цитоплазматическая рибосома состоит из двух субъединиц следующего состава. Малая субъединица — одна длинная цепь рРНК (около 2000 нуклеотидов, константа
седиментации — 18S), с которой связано примерно 30 молекул рибосомальных белков.
Большая субъединица — еще более длинная цепь рРНК (~ 4000 нуклеотидов, 28S), с которой связаны 2 короткие цепи РНК (5,8S и 5S) и около 45 молекул белков.
В итоге каждая субъединица представляет собой свернутый рибонуклеопротеидный тяж, имеющий несколько функциональных центров.
Рибосомные РНК (рРНК) образуются в ядрышках. Там же, в ядрышках, формируются и сами субъединицы, которые затем перемещаются из ядра в цитоплазму.
И в неработающем состоянии субъединицы рибосом так и остаются диссоциированными друг от друга.
Функция рибосом — участие в синтезе белка.
С помощью рибосом осуществляется трансляция — поочередное включение аминокислот в строящуюся пептидную цепь в соответствии с последовательностью кодонов в матричной РНК (мРНК).
! Инициация трансляции.
Для выполнения указанной функции вначале происходит сборка функционально активного комплекса сборка ФАК (что обозначается как инициация трансляции).
5

1.Прежде всего, с определенным центром малой субъединицы связывается начальный участок мРНК.
2.Затем к этому комплексу присоединяется инициирующая транспортная РНК (тРНК), нагруженная первой аминокислотой будущей пептидной цепи.
3.И только после этого, наконец, связывается большая субъединица рибосомы.
!Элонгация и образование полисомы. Собранная рибосома начинает постепенно
перемещаться вдоль мРНК, осуществляя процесс трансляции, т.е. удлиняя строящуюся пептидную цепь. Данная стадия называется элонгацией (отelongation — удлинение).
Когда рибосома удалится на достаточное расстояние от начального участка мРНК, с этим участком связываются субъединицы другой рибосомы (в той же очередности) — так что одну цепь мРНК начинают транслировать сразу две рибосомы.
И так далее: с цепью мРНК постепенно связывается несколько рибосом. Находясь на примерно равном расстоянии друг от друга, они движутся по мРНК в одном направлении, и каждая синтезирует «свою» пептидную цепь. Чем дальше от начала мРНК продвинулась рибосома, тем более длинную пептидную цепь она успела построить.
Такие структуры называются полисомами. Полисома — это комплекс, включающий:
●несколько рибосом
●транслируемую ими цепь мРНК
●несколько (по числу рибосом) пептидных цепей, находящихся на разной стадии синтеза. Состав рибосом в полисоме постепенно сменяется: рибосомы, заканчивающие трансляцию мРНК,
покидают полисому, вновь распадаясь на субъединицы (одновременно высвобождается и готовая
пептидная цепь); а с начальным участком мРНК связываются субъединицы иных рибосом (до тех пор, пока мРНК не будет разрушена специальными РНКазами).
Полисомы
Таким образом, термины «свободные» и «мембраносвязанные» следовало бы применять не к отдельным рибосомам, а к полисомам. Действительно, рибосомы в свободном состоянии практически не существуют: когда они не транслируют какую-либо мРНК, они диссоциированы на отдельные субъединицы.
Полисомы становятся «мембраносвязанными» при образовании экспортных, мембранных, лизосомных и ряда пероксисомных белков, причем с мембраной связывается каждая рибосома полисомы — как только в синтезируемой ею пептидной цепи образуется сигнальная последовательность (СП).
Свободные полисомы синтезируют белки, которые либо остаются в гиалоплазме, либо переходят в состав тех или иных клеточных структур (ядра, митохондрий, цитоскелета, пероксисом). Содержание свободных полисом (а в их составе — и рибосом) особенно велико в быстрорастущих клетках. На свободных полисомах образуются белки, используемые в жизнедеятельности самой клетки, а на прикрепленных — белки, функционирующие вне тела клетки.
6.Эндоплазматическая сеть. Строение, разновидности ЭПС. Структура гранулярной и агранулярной эндоплазматической сети (СМ,ЭМ) и их функции.
6

Эндоплазматическая сеть (ЭПС) - органелла, обеспечивающая синтез углеводов, липидов и белков, а также начальные посттрансляционные изменения
последних. Она имеет мембранное строение.
Состоит из системы уплощенных и удлиненных цистерн, трубочек и везикул, образующих 3-мерную сеть.
Выделяют две разновидности ЭПС: гранулярную ЭПС (грЭПС) и агранулярную ЭПС (аЭПС), которые связаны
друг с другом переходной ЭПС.
В области переходной ЭПС трубочки распадаются на отдельные фрагменты, образующие
окаймленные транспортные пузырьки, которые переносят материал из ЭПС в комплекс Гольджи.
Развитие ЭПС и преобладание одного из видов зависит от типа клетки и её функции.
Гранулярная ЭПС обеспечивает:
●биосинтез всех мембранных белков и белков, предназначенных для экспорта из клетки,
●начальные этапы модификации белковых молекул.
Гранулярная ЭПС образована уплощенными мембранными цистернами и трубочками, на наружной поверхности которых располагаются рибосомы и полисомы.
Мембраны грЭПС содержат особые белки (рибофорины), которые обеспечивают связывание рибосом и уплощение цистерн.
Полость грЭПС содержит продукты синтеза и сообщается с перинуклеарным пространством.
Благодаря грЭПС происходит (изоляция) вновь синтезированных белковых молекул от гиалоплазмы.
Агранулярная (гладкая) ЭПС представляет собой трехмерную замкнутую сеть мембранных
анастомозирующих трубочек, канальцев, цистерн и пузырьков диаметром 20-100 нм. Функции:
4)Синтез липидов, в том числе мембранных
5)Синтез гликогена, холестерина
6)Детоксикация эндогенных и экзогенных веществ
7)Накопление ионов Са2+
8)Восстановление кариолеммы в телофазе митоза.
7.Комплекс Гольджи, (СМ и ЭМ). Полярность комплекса Гольджи. Особенности процессинга молекул и направленный транспорт веществ.
Комплекс Гольджи - сложно организованная мембранная органелла, образованная тремя
основными элементами:
●Цистерны – имеют вид изогнутых дисков и образуют стопку из 3-30 элементов. Выпуклой стороной стопка обычно обращена к ядру, вогнутой - к плазмолемме. Каждая группа цистерн внутри стопки отличается особым составом ферментов, определяющим характер реакций процессинга белков. Периферические отделы цистерн (люмены) несколько расширены,
от них отщепляются пузырьки и вакуоли.
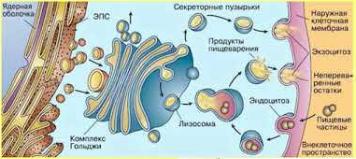
●Пузырьки - сферические окруженные мембраной элементы диаметром 40-80 нм с содержимым умеренной плотности; образуются путем отщепления от цистерн незрелой части.
●Вакуоли - крупные, окруженные мембраной сферические образования, отделяющиеся от цистерны на зрелой поверхности Комплекса Гольджи. Они содержат секреторный продукт
умеренной плотности.
Вспециализированных секреторных клетках комплекс Гольджи располагается надъядерно под апикальной частью клетки, через которую происходит выделение секрета механизмом экзоцитоза. Нередко он лежит у ядра вблизи центриолей, в некоторых клетках его компоненты рассеяны по всей цитоплазме.
Полярность комплекса Гольджи. Комплекс Гольджи представляет собой поляризованную структуру, в которой выделяют две поверхности, обладающие структурными и функциональными различиями:
•Цис-поверхность – незрелую, обращенную к ЭПС и связанную с системой мелких пузырьков, отщепляющихся от ЭПС.
•Транс-поверхность – зрелую, обращенную к плазмолемме и связанную с отделяющимися от цистерн вакуолями
Функции комплекса Гольджи:
1.Синтез полисахаридов и гликопротеинов
2.Процессинг молекул
3.Накопление секреторного продукта и образование секреторных гранул
4.Обеспечение новообразованных гранул мембраной
5.Сортировка белков на транс-поверхности перед их окончательным транспортом
Особенности процессинга молекул.
Входе процессинга в Комплексе Гольджи происходит:
●включение углеводных компонентов в гликопротеины (терминальное гликозилирование)
●добавление фосфатных групп (фосфорилирование),
●добавление жирных кислот (ацилирование),
●добавление сульфатных остатков (сульфатирование),
●частичное расщепление белковых молекул (протеолитическая доработка).
Каждый из указанных этапов процессинга веществ внутри комплекса Гольджи осуществляется в топографически определенном его компоненте (цис-, медиальных или транс-цистернах).
Транспорт белков из Комплекса Гольджи тесно связан с их сортировкой. Сортировка производится посредством специфических мембранных рецепторных белков, которые распознают сигнальные участки на макромолекулах и направляют их в соответствующие пузырьки. Транспорт белков из КГ осуществляется в составе трех важнейших потоков:
9)В гидролазные пузырьки (первичные лизосомы)
10)В плазмолемму
11)В секреторные гранулы
8
8.Структура и функции эндосом и лизосом. Типы эндосом и лизосом
Лизосомы – это разнообразный класс вакуолей размером 0,2 – 0,4 мкм, ограниченных мембраной, являются производным КГ. Характерным признаком лизосом является наличие в них гидролитических ферментов – гидролаз, расщепляющих различные биополимеры при кислом рН.
Основная функция – внутриклеточное переваривание.
Эндосомы - мембранные пузырьки с постепенно закисляющимся содержимым, которые обеспечивают перенос макромолекул с поверхности клетки в лизосомы и их частичный или полный гидролиз на стадиях, предшествующих лизосомальному уровню деградации. Основные функции – транспорт веществ и начальное внутриклеточное переваривание.
Лизосомы и эндосомы представляют собой компартменты (то есть обособленные от цитоплазмы участки с собственной средой для протекания определенных реакций). Создание кислой среды в
этих пузырьках осуществляется за счёт больного количества АТФазных протонных насосов в их мембране.
Также мембрана:
12)Содержит рецепторы, обуславливающие её связывание с мембраной гидролазных и транспортных пузырьков, а также фагосом
13)Обеспечивает свободную диффузию низкомолекулярных продуктов переваривания в гиалоплазму
14)В неповрежденном состоянии представляет собой барьер, резистентный к действию литических ферментов и препятствующий их утечке в гиалоплазму.
Типы эндосом:
!Ранние (периферические) эндосомы – являются мембранными пузырьками на ранних этапах после их отделения от плазмолеммы (но уже после утраты первоначально имевшейся клатриновой оболочки). Они располагаются неподалеку от плазмолеммы в
периферических отделах цитоплазмы. В них происходит:
1)В условиях слабокислой среды (pH 6.0) осуществляется ограниченное и регулируемое переваривание макромолекул протеазами
2)отщепление лигандов от рецепторов с их сортировкой (сортировкой рецепторов) и
возможным возвращением последних в специальных пузырьках в плазмолемму для повторного цикла их использования.
3)расщепление комплексов рецептор-гормон, рецептор-фактор роста, антиген-антитело, а также ограниченный протеолиз (процессинг) антигенов (например, в макрофагах)
!Поздние (перинуклеарные) эндосомы – образуются из ранних и располагаются в глубоких отделах цитоплазмы вблизи ядра. Их отличает от ранних эндосом более плотный матрикс,
более кислое содержимое (pH 5.5) и более глубокий уровень переваривания ферментами.
Типы лизосом:
!Гидролазные пузырьки – округлые мембранные органеллы диаметром до 200-400 нм с мелкозернистым плотным матриксом, содержащие литические ферменты в неактивной форме.
Их перемещение в цитоплазме контролируется микротрубочками. Гидролазные пузырьки участвуют в транспорте литических ферментов в эндоцитозный путь из КГ. В настоящее время известно около 60 литических ферментов; все они представляют собой кислые гидролазы с оптимумом рН~5.
!Лизосомы – органеллы, активно участвующие в завершающих этапах процесса внутриклеточного переваривания захваченных клеткой макромолекул посредством широкого
спектра литических ферментов при низких значениях pH.
9

Их разделяют в зависимости от наличия в их просвете морфологически распознаваемого материала:
●Фаголизосома формируется путем слияния поздней эндосомы или лизосомы с фагосомой.
●Аутофаголизосома образуется при слиянии поздней эндосомы или лизосомы с
аутофагосомой
Мультивезикулярное тельце – крупная сферическая окруженная мембраной вакуоль,
содержащая мелкие пузырьки. Мелкие пузырьки формируются, путем отпочковывания внутрь от мембраны вакуоли. Матрикс тельца содержит литические фермента и, обеспечивает постепенное разрушение внутренних пузырьков.
Остаточные тельца - лизосомы, содержащие непереваренный материал, которые могут
длительно находиться в цитоплазме или выделять свое содержимое за пределы клетки.
9.Митохондрии (СМ и ЭМ). Функции митохондрий
Митохондрии представляют собой двумембранные полуавтономные органеллы, обеспечивающие клетку энергией, получаемой благодаря процессам окисления и запасаемой в виде фосфатных связей АТФ.
Митохондрии также участвуют в:
●биосинтезе стероидов, ●окислении жирных кислот ●синтезе нуклеиновых кислот
Митохондрии могут иметь эллиптическую, сферическую, палочковидную, нитевидную формы. В
цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонем спермия, ресничек), компонентов синтетического аппарата (цистерн ЭПС).
Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны – кристы.
Наружная митохондриальная мембрана напоминает плазмолемму. Она содержит много молекул специализированных транспортных белков (например, порин), которые формируют широкие
гидрофильные каналы и обеспечивают ее высокую проницаемость, а также небольшое количество ферментных систем. На ней находятся рецепторы, распознающие белки, которые переносятся через обе митохондриальные мембраны в особых точках их контакта – зонах слипания.
Внутренняя митохондриальная мембрана отделена от наружной межмембранным пространством. Низкая проницаемость внутренней мембраны для мелких ионов имеет большое значение для функции митохондрий, так как она обеспечивает возможность создания электрохимического градиента.
Всостав мембраны входят:
15)Транспортные белки
16)Ферменты дыхательной цепи и сукцинатдегидрогеназаназа
17)Комплекс АТФ-синтетазы
10