

Лекции УГМУ / Патофизиология нейроэндокринной системы
.pdfмембраны, и что реакция клеток эффекторов реализуется через «циклазную» систему. Циклический 3,5-АМФ образуется из АТФ под влиянием аденилатциклазы, а инактивируется фосфодиэстеразой. Гормоны взаимодействуют с локализованными на внешней поверхности плазматической мембраны клеток рецепторами, сигнал об этом взаимодействии передается на аденилатциклазу, локализованную на внутренней поверхности мембраны, или на фосфодиэстеразу. При этом увеличивается или уменьшается образование циклического 3,5-АМФ, который, в свою очередь, активирует внутриклеточные ферменты. Таким образом, это вещество оказывается как бы внутриклеточным медиатором, который обеспечивает передачу влияния гормона на внутриклеточные ферментные системы. Характер же конечной реакции эффектора определяется спецификой ферментного набора реагирующих клеток, а действие гормона фактически оказывается толчком, пускающим в ход реакцию, предопределенную ферментативным профилем клетки.
Одни клетки, например паренхиматозные клетки печени, обладают рецепторами для малого числа гомонов (только для глюкагона и адреналина). Другие клетки, например адипоциты, имеют рецепторы для большого числа гормонов (глюкагона, секретина, АКТГ, адреналина). Все рецепторы строго специфичны и реагируют только на «свои» гормоны. Инактивация рецепторов для одного из гормонов не влияет на взаимодействие остальных рецепторов с соответствующими гормонами. С другой стороны, представляется весьма вероятным, что все эти рецепторы воздействуют на одну и ту же аденилатциклазу.
Механизмы, которыми амины, гормоны и фармакологические соединения изменяют уровень 3,5-АМФ в разных клетках, не являются идентичными. Увеличение 3,5-АМФ может быть обусловлено активированием циклазы или торможением фосфодиэстеразы.
Из гормонов стимулирующий эффект на биосинтез 3,5-АМФ оказывают: глюкагон, АКТГ, СТГ, ЛГ, антидиуретический гормон, паратиреоидный гормон, меланоцитостимулирующий гормон.
Тормозят фосфодиэстеразу теофиллин, кофеин, папаверин.
11

Рис. 111
Механизм действия нестероидных гормонов (K.I. Catt, M,I. Duffn,
1982)
Участие 3,5-АМФ в обменных процессах осуществляется или активированием протеин-фосфокиназ, или же регуляцией обмена внутриклеточного кальция. Кальций необходим для запуска реакции, которая в дальнейшем стимулируется 3,5-АМФ, или, наоборот, кальций стимулирует реакцию, начатую при участии 3,5-АМФ.
Усиление продукции 3,5-АМФ обусловлено активированием аденилатциклазы, а увеличение концентрации кальция в цитозоле − торможением кальциевых насосов, регулирующих его транспорт из окружающей среды в клетку или же субклеточные образования (кальция вне клетки в 100 раз больше, чем в клетке. Это обеспечивается насосом. Кроме того, установлено, что саркоплазматический ретикулум клетки активно поглощает кальций. Для осуществления обоих этих процессов требуется АТФ). Известно также, что роль рецептора кальция в клетке выполняет белок − тропин. При активации фосфорилирования белков (под влиянием цАМФ) изменяется связь кальция с белком, образуется ионная форма кальция.
12

Взаимосвязь 3,5-АМФ с кальцием можно представить по следующей схеме, представленной на рисунке 112.
Рис. 112
Взаимосвязь 3,5-АМФ с кальцием
Первичный сигнал приводит к активации циклазы и увеличению продукции 3,5-АМФ. Одновременно происходит торможение кальциевого насоса и обогащение внутриклеточного пространства кальцием. Кальций начинает активировать энзиматические системы, связанные с переносом терминального фосфата АТФ на протеин-фосфокиназы. Дальше в обмен кальция вовлекается саркоплазматический ретикулум и сократительный аппарат. В то же время с фосфорилированием связаны в клетке такие процессы, как деятельность натриевого насоса, транспорт и мобилизация ионов кальция.
Высказано предположение, что 3,5-АМФ путем обратного фосфорилирования гистонов вовлекается в изменения генетической экспрессии ДНК.
Следует учитывать, что ионы кальция необходимы и для перевода АТФ в 3,5-АМФ, а также для его распада с образованием 5-АМФ. Аденилатциклаза и фосфодиэстераза свою каталитическую активность проявляют только в присутствии ионов кальция, а также ионов магния.
Определенную роль в реакции аденилатциклазы могут играть и ионы фтора. Экспериментально было установлено, что фториды стимулируют аденилатциклазу. Однако они, в отличие от гормонов, действующих через рецептор на мембране клетки, оказывают прямое действие на аденилатциклазу. Поэтому введение фторидов приводит к тому, что они выигрывают конкурентную борьбу за аденилатциклазу с гормонами и подавляют эффект гормонов на аденилатциклазу.
В передаче импульса с рецептора гормона на аденилатциклазу может играть и другой циклический нуклеотид − гуанозин- 3,5-монофосфат
13

(3,5-ГМФ). Этот нуклеотид участвует во взаимодействии гормона с рецептором и в передаче сигнала от образованного ими комплекса на аденилатциклазу.
Простагландины и 3,5-АМФ. Нервный импульс или гормон инициируют усиление освобождения простагландинов, которые, со своей стороны, могут модифицировать посредством 3,5-АМФ ответную реакцию клетки. Опыты, в которых изучалось действие экзогенных простагландинов, показали, что эти соединения участвуют в функциональной активности клетки посредством 3,5-АМФ. Простагландины находятся в клетке в неактивном состоянии в виде эстеров, их освобождение из связанного состояния стимулируется липолитическими (норадреналин) и тормозится антилиполитическими (инсулин) агентами. Таким образом, в зависимости от условий среды простагландины или действуют на аденилатциклазу, или сами подвергаются воздействию 3,5-АМФ.
Смысл взаимоотношения простагландинов и 3,5-АМФ наиболее четко можно продемонстрировать на примере жировой ткани. Из экспериментальных данных следует, что в ответ на введение адреналина, глюкагона, АКТГ или какого-либо другого липолитического фактора наряду со значительным увеличением синтеза цАМФ в адипоцитах отмечается практически одновременное усиление продукции простагландинов (ПГ) (главным образом типа Е), что ограничивает (возможно, через торможение аденилатциклазы) действие гормона (рис. 113).
Рис. 113
Взаимоотношение простагландинов и цАМФ
Механизм действия стероидных гормонов (рис. 114). Циркулируя в
14

крови, стероиды присутствуют в виде свободных и связанных форм, образующих комплексы с белками плазмы. Основная масса циркулирующих гормонов транспортируется в виде стероидбелковых комплексов. С белками плазмы связано более 98% половых стероидов, около 90% содержащегося в крови кортизола и около 50% альдостерона. Поскольку на уровне клеток-эффекторов активны именно свободные гормоны, роль связывания в плазме может заключаться в создании резервуара или буфера, который контролирует доступность гормона для рецепторов клеток-эффекторов. Поступление стероидов в клетку не зависит от мембранных барьеров или процессов транспорта. Стероиды всех классов вначале связываются со специфическими цитоплазматическими рецепторами, которые подвергаются активации, и комплекс «рецептор-гормон» перемещается из цитоплазмы в ядро. Здесь происходит связывание комплекса ядерными акцепторными участками хроматина клеток-эффекторов, что модулирует активность специфических генов, ответственных за синтез определенных видов мРНК. Процесс активации рецептора индуцируется гормоном и зависит от температуры.
Действие гормонов может проявляться и качественными изменениями ферментных «наборов» реагирующих клеток, что приводит к качественным изменениям метаболизма, к дифференцировке сперва биохимической, а затем (по достижении достаточной интенсивности) и к структурной, то есть к морфогенетическому эффекту.
Рис. 114
Механизм действия стероидных гормонов (K.I. Catt, M.I. Dufan,
1982)
C – стероид, Р – рецептор. Активация внутриклеточных рецепторов
15
стероидными гормонами сменяется ядерным связыванием комплекса и стимуляцией синтеза мРНК.
Особое место занимают тиреоидные гормоны.
Механизм действия тиреоидных гормонов (рис. 115). Тиреоидные гормоны оказывают многочисленные и разнообразные эффекты на дифференцировку, развитие и метаболический гомеостаз, контролируя синтез
иактивность регуляторных белков, в том числе ключевых ферментов метаболизма, гормонов и рецепторов. Известное действие тиреоидных гормонов на потребление кислорода определяется отчасти стимуляцией натриевого насоса за счет индукции мембранного фермента натрий-калий-зависимой АТФазы. Этот и другие метаболические эффекты тиреоидных гормонов зависят от гормональной индукции синтеза РНК, осуществляемой путем регуляции экспрессии генов на ядерном уровне. Хотя существует и прямое влияние тиреоидных гормонов на клеточную мембрану
ина митохондрии.
Основным йодтиронином, секретируемым щитовидной железой, является тироксин (Т4), которому сопутствует небольшое количество трийодтиронина (Т3). В клетках-эффекторах Т4 дейодируется в Т3, который представляет собой главную внутриклеточную форму гормона. Это превращение происходит в плазматической мембране и эндоплазматическом ретикулуме. Хотя и показано присутствие связывающих Т3 белков в цитоплазме, но они обладают относительно низким сродством к гормону. Поэтому в качестве истинного рецептора Т3 рассматриваются ядерные участки, ответственные за эффекты тиреоидных гормонов.
16
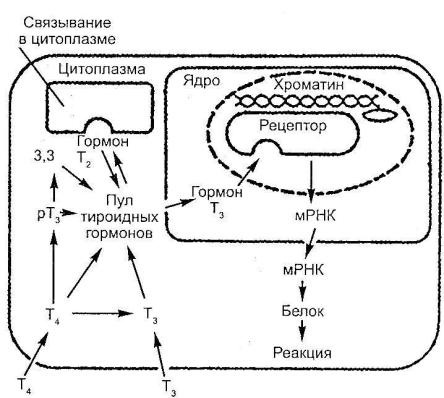
Рис. 115
Механизм действия тиреоидных гормонов (K.I. Catt, M.I. Dufan,
1982)
Эффекты и взаимоотношение Т3 и Т4 в клетках, реагирующих на поступление тиреоидных гормонов. Процесс активации ядра зависит от связывания Т3 с ядерными рецепторами, а не от отдельно существующих цитоплазматических рецепторов, как это имеет место в клетках-мишенях стероидных гормонов.
Строение и функция эндокринной системы
Центром эндокринной системы принято считать гипоталамо-гипофизарный комплекс. Прочие железы внутренней секреции обозначают как периферические. Это разделение основано на том, что в числе гормонов, продуцируемых передней долей гипофиза, вырабатываются так называемые тропные или кринотропные, которые активируют некоторые периферические эндокринные железы.
На основании морфологических и функциональных критериев этот комплекс можно разделить на две части:
1. Систему, состоящую из гипоталамуса и нейрогипофиза (задней доли гипофиза) − в нервных клетках супраоптического и паравентрикулярного ядер вырабатываются АДГ (антидиуретический гормон) и окситоцин, которые по аксонам нейросекреторных клеток, образующих гипоталамо-гипофизарный тракт, поступают в заднюю долю гипофиза, где терминали аксонов образуют тесные контакты с капиллярами, куда и секретируют гормоны при возбуждении нейросекторных клеток.
17

2. Систему, образованную гипофизотропной зоной гипоталамуса, которая располагается в серединном возвышении, связанной с аденогипофизом (передней долей гипофиза) посредством воротной системы гипофиза (эта воротная система начинается ветвями верхней гипофизарной артерии, которые распадаются на мелкие капиллярные петли, возвратные ветви которых, образуя воротные вены, несут кровь к капиллярному сплетению аденогипофиза). В дополнение к нейросекреторным клеткам, вырабатывающим АДГ и окситоцин, гипоталамус содержит мелкие нервные клетки, открытые позднее первых, которые вырабатывают факторы, стимулирующие или ингибирующие высвобождение гормонов передней доли гипофиза − гипофизотропные гормоны. Так как аденогипофиз, в свою очередь, регулирует секрецию многих других гормонов, эта гипофизарная зона представляет собой узловой участок коммуникационных путей, связывающих нервную систему с эндокринной.
Таким образом, нейросекреторные клетки гипоталамуса являются местом трансформации нервных стимулов в гуморальные факторы, обладающие высокой специфичностью и физиологической активностью. Они, владея, по образному выражению Эрнста Шаррера (E. Scharrer) (1952), «двумя языками» − нервным и гуморальным, играют роль своего рода переводчиков «языка приказаний нервной системы на язык гуморальной активности».
Гипоталамус структурно и функционально связан со средним мозгом, лимбической системой и ринэнцефалическими ядрами переднего мозга.
Влияние гипоталамуса на гипофиз осуществляется по меньшей мере тремя путями: нервные механизмы; секреция гипофизотропных гормонов (релизинг-факторов); через пептиды − энкефалины и эндорфины − так называемые «эндогенные опиаты» (наиболее характерный их признак − морфиноподобное действие), нейротензин, вещество Р и т.д. Роль последней группы веществ в настоящее время окончательно не выяснена в механизмах эндокринной регуляции. Однако, поскольку общей их особенностью является гипофизотропная активность, то все они родственны релизиг-факторам.
Учитывая, что ряд из этих веществ − субстанция Р (угнетает стимулированную кортикотропин-релизинг-фактором секрецию АКТГ), нейротензин (принимает участие в стимуляции секреции СТГ, пролактина, ТТГ и АКТГ, гонадотропных гормонов) − вырабатывается в желудочно-кишечном тракте клетками, входящими в систему APUD, следует предположить, что функция гипофиза контролируется не только гипоталамусом, но на нее могут влиять и другие отделы эндокринной системы.
Все периферические железы внутренней секреции можно разделить на две основные группы по отношению к передней доле гипофиза.
Первую группу составляют гипофиззависимые железы: щитовидная железа, гонады и кора надпочечников (глюкокортикоидная функция
18
надпочечников). Зависимость этих желез от передней доли гипофиза настолько велика, что гипофизэктомия влечет за собой резкое ослабление их функциональной деятельности и даже атрофию их паренхимы. В свою очередь, гормоны гипофиззависимых периферических желез действуют на переднюю долю гипофиза, угнетая продукцию и секрецию соответствующих кринотропных гормонов. Таким образом, реципрокные взаимоотношения между передней долей гипофиза и активируемыми ею периферическими железами имеют характер отрицательных обратных связей, или «плюс-минус взаимодействий» (как их в свое время обозначали).
Вторую группу периферических желез внутренней секреции составляют независимые от передней доли гипофиза железы. К группе независимых принадлежат околощитовидные железы, клубочковая зона коры надпочечников, продуцирующая минералокортикоиды, хромаффинные клетки мозговой части надпочечников, К-клетки щитовидной железы, островки Лангерганса поджелудочной железы, энтерохромаффинные и аргирофильные клетки желудка и кишечника. Инкреторная активность этих желез определяется непосредственно уровнем их гормонов, находящихся в циркуляции, или интенсивностью тех эффектов, которые вызываются в организме гормонами этих желез. Поэтому независимые железы могут рассматриваться как саморегулирующиеся. Однако выраженная саморегуляция деятельности этих желез отнюдь не означает их автономности. Из того, что регуляция некоторых периферических эндокринных желез обеспечивается без обязательного участия кринотропных гормонов гипофиза, вовсе не следует, что они вообще не испытывают зависимости от организма в целом. В частности, в некоторых из таких желез достаточно отчетливо обнаруживается зависимость от прямых нервных импульсов (например, мозговой слой надпочечников, эндокринная часть поджелудочной железы). Кроме того, СТГ, влияя на синтез белков, может оказывать влияние и на образование белкового гормона − инсулина в поджелудочной железе.
Сдругой стороны, способность к саморегуляции иногда проявляется и
ужелез, деятельность которых явно активируется гипофизарными кринотропными гормонами.
Следует отметить, что эндокринные железы, удаление которых неизбежно приводит к смерти (околощитовидные, клубочковая зона коры надпочечников, панкреатические островки), относятся к группе саморегулирующихся желез.
Экскреция же зависимых желез и даже самого гипофиза не является смертельной, и жизнь без них может продолжаться, хотя и с существенными отклонениями от нормы.
Необходимо учитывать, что взаимодействие между эндокринными органами далеко не исчерпывается реципрокными связями между периферическими железами и передней долей гипофиза. Не исключена возможность действия гормона одной железы на другую не только
19
опосредованно через переднюю долю гипофиза, но и непосредственно (например, тиреоидных гормонов на островки Лангерганса или на кору надпочечников, инсулина на щитовидную железу и т.д.). Влияние может осуществляться помимо прямого действия гормонов и в результате изменения эффектов, вызываемых в организме гормонами гипофиззависимых желез. Например, щитовидная железа или кора надпочечников могут оказывать влияние на поджелудочную железу не столько непосредственно своими гормонами, сколько тем, что эти гормоны участвуют в контролировании углеводного обмена в организме, изменяя уровень сахара в крови или содержание гликогена в печени, что, в свою очередь, сопровождается сдвигами в секреции инсулина и глюкагона вследствие отмеченного изменения уровня сахара в крови.
Наконец, разбросанные по разным органам и тканям клетки, вырабатывающие регуляторные пептиды (субстанцию Р, нейротензин, соматостатин, холецистокинин, вазоактивный интестинальный пептид [ВИП]) и амины (серотонин, гистамин, кинины), образуют диффузную эндокринную систему − систему APUD, связанную с интегральной эндокринной системой, представленной перечисленными выше структурами (по крайней мере, на уровне гипофиза).
Кроме уже названных структур, образующих эндокринную систему, следует еще упомянуть юкстагломерулярный аппарат (ЮГА) почки, работающий как эндокринная железа и выделяющий по меньшей мере два вещества, относимых исследователями к гормонам: ренин и эритропоэтин.
В предсердиях вырабатывается увеличивающий почечную фильтрацию, уменьшающий канальцевую реабсорбцию, снижающий секрецию почками ренина гормон − атриопептин (синонимы: натрийуретический фактор − ANF [atrium natriuretic factor], аурикулин, кардинатрин).
Образуются вещества с гормональной активностью в кишечнике. Это гастрин, холецистокинин.
Все без исключения железы внутренней секреции находятся под постоянным контролем вегетативной нервной системы − ее симпатического и парасимпатического отделов. Неоднократно высказывалось мнение, что нервы эндокринных желез выполняют главным образом сосудодвигательную функцию. В этом случае кровеносные сосуды (их тонус, скорость кровотока) являются основным рычагом, посредством которого эфферентные нервы оказывают влияние на гормонопоэз. Кроме того, известно, что медиаторы симпатических и, возможно, парасимпатических окончаний нервов кровеносных сосудов могут частично выделяться в межклеточную жидкость и оказывать прямое влияние на паренхиматозные клетки, регулируя уровень усвоения этой клеткой питательных веществ и чувствительность к гуморальным раздражителям (релизинг-факторам гипоталамуса, кринотропным гормонам гипофиза, кортикостероидам, глюкозе, кальцию
20