

11. Прессорный диурез. Предсердный натрийуретический гормон — место выработки, стимуляторы секреции, эффекты, механизмы действия.
Рассмотрим последовательность реагирования организма на изменения солевой нагрузки.
При повышении потребления NaCl: кровь становится гиперосмолярной; стимуляция осморецепторов гипоталамуса вызывает выброс АДГ; увеличивается реабсорбция воды;
кровь становится нормоосмолярной, но возрастает ее объем (развивается гиперволемия);
гиперволемия запускает системы, повышающие выведение натрия и воды (натрийурез и диурез), и подавляет системы, снижающие натрийурез и диурез; избыток изотоничной жидкости выводится с мочой.
При снижении потребления NaCl развиваются обратные процессы.
Итак, изменения солевой нагрузки компенсируются в два этапа.
1. Включается система АДГ, нормализующая осмолярность крови, но ценой изменений ее объема.
2. Измененный объем крови корректируется системами регуляции натрийуреза (и одновременно диуреза). Все эти системы, как уже говорилось, регулируют выведение примерно изотоничной мочи.
Предсердный натрийуретический гормон
Этот гормон вырабатывается в предсердиях (секреторными кардиомиоцитами) в ответ на их растяжение повышенным объемом крови; вызывает уменьшение реабсорбции Nа+ в корковых отделах собирательных трубочек и тем самым усиленное выделение Nа+ (натрийурез) и воды (диурез). Является антагонистом деятельности ренин-ангиотензиновой системы (тормозит секрецию ренина и снижает прессорную активность ангиотензина II). Также тормозит активность альдостерона, АДГ; усиливает клубочковую фильтрацию.
Предсердный натрийуретический пептид секретируется в ответ на: растяжение предсердий, гипернатриемию, хотя повышение натрия в крови не является прямым стимулом к высвобождению ПНГ; ангиотензин-II; эндотелин.
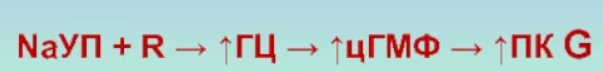
ПК G фосфорилирует Na+, K+-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению насоса. Результат этого – ослабление реабсорбции ионов натрия, а значит и воды.
Прессорный диурез
Это система долгосрочного регулирования, отвечающая за поддержание постоянного уровня среднего артериального давления на протяжении часов, суток и дольше.
Прессорный диурез заключается в том, что при повышении среднего артериального давления почки автоматически увеличивают выведение натрия (натрийурез) и воды. Это приводит к снижению ОЦК и, как следствие, среднего артериального давления.
При длительном повышении артериального давления прессорный натрийурез существенно усиливается, поскольку высокое давление подавляет выделение ренина почкой, вследствие этого снижается выработка ангиотензина II и альдостерона. Низкий уровень ангиотензина II и альдостерона тормозят реабсорбцию натрия в канальцах, усиливая прямой эффект повышенного давления на увеличение выделения натрия и воды с мочой.
12. Почечная регуляция концентрации в крови калия, кальция и фосфата. Механизмы секреции калия. Действие на почки птг и кальцитонина.
Почечная регуляция кислотно-щелочного равновесия
Вещества, имеющие кислую или щелочную реакцию, поступая в кровь, реагируют с бикарбонатным буфером, изменяя соотношение его компонентов — Н2СО3 (СО2) и НСО3-; задача выделительных систем — поддерживать концентрации компонентов бикарбонатного буфера (СО2 и НСО3- или, по крайней мере, соотношение этих концентраций; концентрация СО2 регулируется легкими, а концентрация НСО3- — почками.
Таким образом, роль почек в поддержании кислотно-щелочного равновесия заключается в регуляции концентрации бикарбоната в крови.
Эта роль выполняется следующим образом. За сутки отфильтровывается огромное количество (около 4300 ммоль/сут) бикарбоната, и далее:
1. Если избытка кислых или щелочных веществ в крови нет — отфильтрованный бикарбонат почти полностью реабсорбируется;
2. Если есть избыток щелочных веществ — отфильтрованный бикарбонат не полностью реабсорбируется, то есть частично выводится с мочой (напомним, что бикарбонат — щелочное вещество);
3. Если есть избыток кислых веществ — не только полностью реабсорбируется отфильтрованный бикарбонат, но в почках еще и образуется и выделяется в кровь дополнительное количество бикарбоната — так называемый новый бикарбонат.
Почечный транспорт бикарбоната подчиняется общим принципам обязательного и факультативного транспорта:
1. В проксимальном канальце в обязательном порядке реабсорбируется основная часть (80—90%) отфильтровавшегося бикарбоната, при этом рН канальцевой жидкости меняется мало.
Если бы бикарбонат реабсорбировался в таком же относительном количестве, что и вода, то рН канальцевой жидкости, разумеется, не менялся бы вовсе и оставался бы равным 7,4. Однако вода в проксимальном канальце реабсорбируется на 65% (закон двух третей), а бикарбонат — на 80—90%, поэтому концентрация бикарбоната в канальцевой жидкости уменьшается, а соответственно снижается и рН — до 6,7.
2. В дистальном отделе, в зависимости от кислотно-щелочного состояния организма, происходят процессы, перечисленные выше.
Проксимальный каналец - реабсорбция бикарбоната
Содержащийся в канальцевой жидкости бикарбонат сам по себе не может пройти в клетку эпителия почечного канальца через апикальную мембрану, так как в этой мембране нет переносчиков бикарбоната. В связи с этим общий принцип переноса бикарбоната через апикальную мембрану следующий: бикарбонат в канальцевой жидкости переходит в СО2, последний, будучи жирорастворимым, свободно проходит через апикальную мембрану, а в клетке снова переходит в бикарбонат. Из клетки бикарбонат с помощью переносчика выводится через базолатеральную мембрану в интерстиций. Ключевую роль в этих процессах играют два фактора:
1. Для того, чтобы перейти в СО2, бикарбонат в канальцевой жидкости должен сначала соединиться с протоном (Н+), образующимся в канальцевой клетке из угольной кислоты;
2. Образование и распад угольной кислоты катализируются ферментом карбоангидразой, содержащейся как внутри канальцевых клеток, так и на поверхности их апикальной мембраны (на щеточной каемке).
Последовательность происходящих при этом процессов следующая:
В клетке эпителия почечного канальца из СО2 и воды под действием внутриклеточной карбоангидразы образуется угольная кислота.
Угольная кислота диссоциирует до НСО3- и Н+.
НСО3- выводится совместным переносом с Nа+ через базолатеральную мембрану в интерстиций.
Н+ выводится через апикальную мембрану в канальцевую жидкость Nа+/Н+-обменником (вторичным активным контртранспортом).
В канальцевой жидкости Н+ реагирует с НСО3- с образованием угольной кислоты.
Под действием карбоангидразы щеточной каемки апикальной мембраны угольная кислота распадается до СО2 и воды.
СО2, будучи жирорастворимым, проходит через мембрану.
Вошедший СО2 вновь соединяется с водой, образуя угольную кислоту, и весь цикл повторяется.
Итак, в проксимальном канальце:
1. Бикарбонат проникает из канальца в клетку, а из клетки переносится в интерстиций — иными словами, происходит реабсорбция бикарбоната;
2. Н+ выходит из клетки в каналец, но затем, прореагировав с бикарбонатом, обратно входит в нее в составе воды — иными словами, в целом не происходит секреции протонов, но Н+ служит лишь своего рода челноком для переноса бикарбоната;
3. Для сохранения электронейтральности вместе с бикарбонатом реабсорбируется Nа+: он переходит из канальца в клетку с помощью Nа+/Н+-обменника, а затем выходит из клетки совместным переносом с бикарбонатом.
Дистальный извитой каналец - образование нового бикарбоната
Наблюдается явное сходство механизмов реабсорбции бикарбоната и образования нового бикарбоната. В обоих случаях в клетке под действием карбоангидразы образуется угольная кислота, которая затем распадается до Н+, выходящего в канальцевую жидкость, и НСО3- , выходящего в интерстиций. Главное же отличие заключается в том, что при образовании нового бикарбоната выходящий в канальцевую жидкость Н+ не соединяется с НСО3- и не входит с ним обратно в клетку в составе СО2 и воды. Это связано с тем, что в жидкости дистального извитого канальца (где происходит образование нового бикарбоната) НСО3- почти нет — он почти полностью реабсорбировался в предыдущих отделах нефрона.
Последовательность происходящих при образовании нового бикарбоната процессов следующая:
В клетке из СО2 (поступающего не из канальца, а из крови!) и воды под действием карбоангидразы образуется угольная кислота.
Угольная кислота диссоциирует до бикарбоната и Н+.
Бикарбонат выводится через базолатеральную мембрану в интерстиций хлор-бикарбонатным обменником (вторичным активным контртранспортом).
Н+ выводится через апикальную мембрану в канальцевую жидкость Н+-АТФазой (протонным насосом).
Вошедший из интерстиция в обмен на бикарбонат хлор выводится в каналец через хлорные каналы.
Итак, в дистальном извитом канальце:
1. Бикарбонат, выходящий в интерстиций, не реабсорбируется из канальцевой жидкости, но образуется в канальцевой клетке из поступающего из крови СО2 — иными словами, происходит образование нового бикарбоната;
2. Н+ выходит из клетки в каналец и остается в канальцевой жидкости, закисляя ее — иными словами, происходит секреция протонов;
3. Для сохранения электронейтральности происходит секреция хлора: в базолатеральной мембране он обменивается на бикарбонат, а в апикальной выходит в канальцевую жидкость вслед за протоном.
Важно, что выделяющийся в дистальный извитой каналец Н+ не пребывает в канальцевой жидкости в свободном виде, но связывается с буферами мочи — фосфатным и аммиачным. Фосфатный буфер состоит из сравнительно небольшого числа отфильтровавшихся и не реабсорбированных в проксимальном отделе канальцев фосфатов. Аммиачный буфер представлен аммиаком, образующимся в канальцевых клетках в процессе распада глутамина. Из этих двух буферов самый мощный — аммиачный. Если бы этих буферов не было, то для выведения даже того небольшого количества протонов, которое в норме вырабатывается ежесуточно в составе нелетучих кислот, потребовалось бы выделять либо чрезвычайно кислую мочу, либо огромное ее количество.
Приведем примерные расчеты. За сутки вырабатывается 40—90 ммоль (в пересчете на Н+) нелетучих кислот (в основном продукта распада серосодержащих аминокислот — серной кислоты), а значит, должно выводиться 40—90 ммоль протонов. Если округленно принять суточную выработку нелетучих кислот равной 100 ммоль, а суточное выделение мочи — 1 л, то концентрация протонов в моче в отсутствие буферов составила бы 100 ммоль/л = 0,1 моль/л; это соответствует рН = 1. Такая высокая кислотность не достигается даже в желудке на пике секреции соляной кислоты. На самом деле минимальный рН мочи составляет 4,5, что соответствует концентрации протонов около 0,00003 моль/л (10-4,5 = 0,00003162); значит, для выведения 100 ммоль протонов в отсутствие буферов потребовалось бы более 3000 л мочи в сутки!
Почечная регуляция электролитного состава крови
Калий
Поскольку человеку приходится преимущественно сталкиваться с поступлением большого количества калия с пищей (а не с его чрезмерной потерей), основные механизмы почечной экскреции калия направлены на выведение его избытка (а не на задержку в организме). Это означает, что в почках особую роль играет факультативная секреция калия.
Почечный транспорт калия подчиняется общим принципам обязательного и факультативного транспорта:
1. В проксимальном канальце в обязательном порядке реабсорбируется 2/3 (65%) отфильтровавшегося калия;
2. В дистальном отделе происходит в основном дополнительная секреция калия, интенсивность которой зависит от поступления калия в этот отдел.
Существуют и механизмы факультативной реабсорбции калия, включающиеся при его потерях, однако их роль и мощность гораздо меньше.
Механизмы секреции калия
1. Через базолатеральную мембрану калий переносится в клетку Nа+, К+-АТФазой.
2. Через апикальную мембрану калий выходит в каналец по калиевым каналам.
Регуляция секреции калия
Секреция калия повышается в ответ на гиперкалиемию (увеличение концентрации калия в крови). Это обусловлено в основном (но не исключительно) тем, что гиперкалиемия — один из двух главных стимуляторов (наряду с ангиотензином II) выброса альдостерона корковым веществом надпочечников. Альдостерон путем ряда механизмов повышает секрецию калия (и одновременно — реабсорбцию натрия) в дистальном отделе нефрона; важнейший из этих механизмов — увеличение синтеза Nа+, К+-АТФазы.
Кальций и фосфат
Кальций относится к веществам, подчиняющимся «правилу двух третей»: он на 65% реабсорбируется в проксимальном канальце. ПТГ (паратиреоидный гормон) повышает факультативную реабсорбцию кальция в дистальном отделе нефрона, тем самым уменьшая его выведение с мочой.
Фосфат относится к пороговым веществам: он появляется в моче, когда его концентрация в крови становится выше пороговой (однако пороговая концентрация фосфата, в отличие от глюкозы и аминокислот, не выше нормальной концентрации в крови, поэтому он может присутствовать в моче даже в норме). ПТГ снижает реабсорбцию фосфата, тем самым увеличивается выведение фосфата с мочой. Таким образом, ПТГ уменьшает выведение с мочой кальция, но увеличивает — фосфата.
Кальцитонин – синтезируется «С» клетками щитовидной железы. Секреция инициируется повышением концентрации Ca2+ в крови. Является антагонистом паратгормона. Подавляет реабсорбция Ca2+ в почках, значит, повышает выведение кальция с мочой. Эффект: нормализация концентрации Ca2+ в крови. Также кальцитонин может действовать на костную ткань, тормозя активность остеокластов угнетает процесс резорбции костной ткани.
Кальцитриол (активная форма витамина D) – кожа (холекальциферол) печень (кальцидиол) почки (кальцитриол). Усиливает реабсорбцию Ca2+ и фосфатов в почках, усиливает всасывание Ca2+ и фосфатов в ЖКТ, облегчает выход Ca2+ из костей.
Клиренс, или коэффициент очищения, — важнейший показатель функции почек. По определению, клиренс некоего вещества равен объему плазмы, полностью очищающемуся от данного вещества за единицу времени. Важно подчеркнуть, что этот объем — условный. Например, клиренс инулина равен 180 л/сут; это означает, что почки способны были бы за сутки полностью очистить от этого вещества 180 л плазмы.
Представим себе, что для некоего вещества X концентрация в плазме равна Сп, в моче — См, а суточный диурез равен Д (суточный диурез необходимо перевести из л/сутки в мл/минуту.
![]()