

7. Пороговые вещества. Причина существования пороговой концентрации ряда веществ в крови. Важнейшие пороговые вещества.
Любые виды транспорта, требующие переносчиков (первичный активный, вторичный активный, облегченная диффузия), характеризуются насыщением. Это означает, что при относительно низких концентрациях некоторых веществ в крови (а следовательно, и в фильтрате) переносчики успевают полностью реабсорбировать такие вещества и в окончательной моче их нет. При превышении же некоторой критической, или пороговой, концентрации переносчики становятся полностью занятыми (насыщаются), вещество уже не может реабсорбироваться полностью и появляется в моче. Такие вещества называются пороговыми. К важнейшим из пороговых веществ относятся: глюкоза, аминокислоты, бикарбонат, фосфат.
Для полезных органических субстратов (глюкозы и аминокислот) пороговая концентрация намного выше нормальной концентрации в крови, поэтому в норме эти вещества с мочой не выводятся (они появляются в моче только при патологии, например при сахарном диабете, когда концентрация глюкозы в крови резко возрастает и становится выше пороговой). Для бикарбоната пороговая концентрация лишь ненамного выше нормальной концентрации в крови, поэтому обычно его в моче нет, но он выводится с мочой даже при небольшом защелачивании крови (алкалозе), что имеет первостепенное значение для поддержания кислотно-щелочного равновесия. Наконец, для фосфата пороговая концентрация ниже верхней границы нормы содержания в крови, поэтому он может содержаться в моче даже в обычных условиях, образуя одну из буферных систем мочи — фосфатный буфер (но при низком содержании фосфата в пище, а соответственно и низкой концентрации в крови, он полностью реабсорбируется и в моче не содержится).
Пороговая концентрация для глюкозы равна 10 ммоль/л.
8. Петля Генле, ее основное назначение. Особенности процессов секреции и реабсорбции.
9. Роль дистальных отделов нефрона в регуляции воды и натрия. Механизмы реабсорбции и секреции в дистальных отделах нефрона.
Почечная регуляция водно-осмотического равновесия
Поддержание водно-осмотического равновесия включает два компонента: поддержание объема жидкости в организме; поддержание ее осмолярности.
Для этого необходимо, чтобы: суточное поступление воды (суточная водная нагрузка) соответствовало ее суточному удалению; суточное поступление осмотически активных веществ (суточная осмотическая нагрузка) соответствовало их суточному удалению.
Иногда оба эти требования выполнить невозможно, и тогда приходится выбирать между поддержанием объема жидкости в организме и ее осмолярности.
В таких случаях поддержанию осмолярности отдается более высокий приоритет (например, на поступление такого большого количества соли, с выведением которого почки не справляются, организм реагирует увеличением потребления и (или) снижением выделения воды; объем жидкости в организме при этом возрастает, но осмолярность ее остается прежней).
В идеале почки должны полностью справляться и с суточной водной нагрузкой, и с суточной осмотической нагрузкой. Следовательно, для того чтобы понять требования, предъявляемые к почечным системам регуляции водно-осмотического баланса, надо рассмотреть, какие диапазоны этих нагрузок характерны для среды обитания и образа жизни человека, ведь именно к этим диапазонам в процессе эволюции приспособились регуляторные системы.
1. Суточная водная нагрузка у человека колеблется в диапазоне от 0 до 20 л.
2. Суточная осмотическая нагрузка подразделяется на экзогенную, основную часть которой составляет NaCl (солевая нагрузка), и эндогенную, основную часть которой составляет образующаяся при распаде белков мочевина (мочевинная нагрузка):
суточная солевая нагрузка колеблется от 1 до 10 г NaCl (диапазон потребления соли у человека), то есть от 30 до 300 миллиосмолей.
минимальная суточная мочевинная нагрузка при обычном питании и образе жизни составляет 500 миллиосмолей, что уже больше, чем максимальная суточная солевая нагрузка. С учетом того, что потребление белка, а следовательно, и образование мочевины, может возрастать в несколько раз, очевидно, что суточная мочевинная осмотическая нагрузка существенно больше солевой.
Таким образом, к почечным системам регуляции водно-осмотического равновесия предъявляются следующие требования:
- они должны справляться с очень большой водной нагрузкой, регулируя выведение воды в чрезвычайно широком диапазоне;
- они должны справляться со значительной мочевинной осмотической нагрузкой, также регулируя выведение мочевины в широком диапазоне;
- они должны справляться с существенно меньшей солевой осмотической нагрузкой;
- в случае если все эти требования одновременно удовлетвориться не могут, приоритет должен отдаваться поддержанию осмолярности, а не объема.
Показатели, отражающие функцию почек по поддержанию водно-осмотического баланса, — это суточный диурез (суточный объем мочи) и осмолярность мочи.
Осмолярность мочи может колебаться от 50 до 1200 мосмоль/л (осмолярность крови — в среднем 285 мосмоль/л); иными словами, моча может быть в 6 раз гипотоничнее или в 4 раза гипертоничнее крови.
Предельные значения диуреза определяются этими предельными значениями осмолярности мочи и суточной осмотической нагрузкой.
Средняя суточная осмотическая нагрузка составляет около 600 миллиосмолей (500 миллиосмолей за счет мочевины; остальные 100 миллиосмолей за счет NaCl и некоторых других веществ). Для такой осмотической нагрузки минимальный диурез (при максимально концентрированной моче) составляет 600 (мосмоль/сут)/1200 (мосмоль/л) = 0,5 л; максимальный диурез (при максимально разбавленной моче) составляет 600 (мосмоль/сут)/50 (мосмоль/л) = 12 л. Существенно уменьшить суточную осмотическую нагрузку без ущерба для организма нельзя, так как эта нагрузка создается в основном мочевиной, и поэтому снизить ее можно только за счет уменьшения потребления белка ниже предельно допустимых величин. Увеличить же суточную осмотическую нагрузку в известных пределах можно, и тогда будет возрастать и максимальный диурез, достигая в крайних случаях 20 л/сут. Таким образом, суточный диурез может колебаться от 0,5 до 20 л.
Итак, при полном отсутствии потребления воды выделяется 0,5 л/сут предельно концентрированной мочи (1200 мосмоль/л). При резко повышенном потреблении воды может выделяться до 20 л/сут предельно разбавленной мочи (50 мосмоль/л).
Регуляция выведения воды
Для регуляции выведения воды почки должны уметь выделять как гипертоничную, так и гипотоничную мочу, то есть обладать разбавляющей и концентрирующей функцией.
Механизм разбавления мочи
Из особенностей трансэпителиального переноса видно, что вода всегда реабсорбируется путем осмоса, то есть следует за реабсорбируемыми растворенными веществами, и в основном — за натрием. Следовательно, если в некоем отделе нефрона реабсорбируются растворенные вещества и он высоко проницаем для воды — будет происходить изоосмотическая реабсорбция: и реабсорбируемая, и канальцевая жидкость будут изотоничны. Если же будут реабсорбироваться растворенные вещества, но каналец будет непроницаем для воды — реабсорбируемая жидкость будет гипертонична, а канальцевая — гипотонична.
Как мы уже знаем, проксимальный каналец проницаем для воды, и потому из него оттекает изотоничная жидкость. Напротив, восходящая часть петли Генле и ранний отдел дистального извитого канальца всегда непроницаемы для воды, а NaCl в них реабсорбируется, поэтому от них оттекает гипотоничная жидкость с осмолярностью 100 мосмоль/л.
Реабсорбция NaCl продолжается в позднем отделе дистального извитого канальца и в собирательной трубочке; эти отделы в отсутствие АДГ непроницаемы для воды, и в этих условиях осмолярность мочи в конечном счете может снижаться до минимального значения — 50 мосмоль/л.
Механизм концентрирования мочи
Из предыдущего раздела мы узнали, как почки могут образовывать изотоничную и гипотоничную мочу. Остается выяснить, как почки могут выделять гипертоничную мочу.
Предположим, что через интерстиций рядом друг с другом проходят два канальца. В одном из них в интерстиций реабсорбируются растворенные вещества, но не вода; интерстиций при этом становится гипертоничным. Другой же каналец, наоборот, проницаем для воды, но в нем не реабсорбируются растворенные вещества; тогда вода будет выходить из него по градиенту осмотического давления в интерстиций, и жидкость в канальце станет гипертоничной. Таким образом, принцип концентрирования мочи заключается в том, что одни отделы нефрона путем реабсорбции растворенных веществ создают повышенное осмотическое давление в интерстиции, а из других в этот гипертоничный интерстиций выходит вода.
Отделом, создающим гипертоничность интерстиция, является петля Генле.
Отделом, из которого в этот гипертоничный интерстиций выходит вода (а канальцевая жидкость при этом концентрируется), является собирательная трубочка.
Создание гипертоничной среды в интерстиции резко усиливается за счет работы петли Генле как поворотно-противоточной системы.
Чтобы понять назначение поворотно-противоточной системы, рассмотрим сначала, какова была бы эффективность концентрирования интерстиция в ее отсутствие.
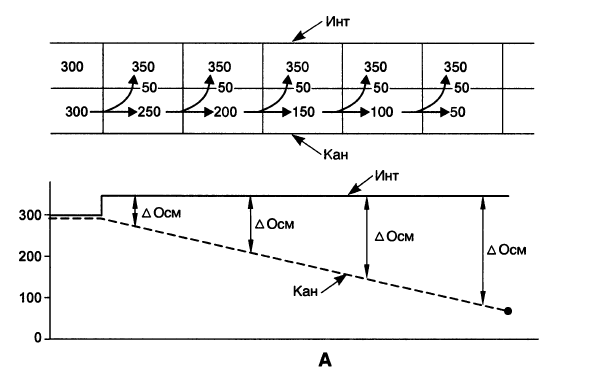
На рис. 15.8, A изображен каналец, проходящий через интерстиций. Предположим, что жидкость в канальце течет в направлении, указанном прямыми стрелками, а интерстициальная жидкость неподвижна. И сам каналец, и интерстиций разделены на несколько условных квадратиков. Пусть изначально осмолярность и канальцевой, и интерстициальной жидкости равна 300 мосмоль/л. Из канальца в интерстиций реабсорбируется NaCl с такой интенсивностью, что в каждом условном квадратике в интерстиций выходит 50 мосмоль/л этого электролита. Тогда канальцевая жидкость по мере продвижения будет становиться все более гипотоничной (на 50 мосмоль/л в каждом квадратике), а интерстиций в каждом квадратике будет становиться гипертоничнее на 50 мосмоль/л; иными словами, осмолярность интерстиция везде станет равной 350 мосмоль/л. Эти процессы отражены на графике в нижней части рис. 15.8, А. Из него видно, что: осмолярность интерстиция выросла на всем его протяжении лишь на 50 мосмоль/л; по ходу канальца создается нарастающий осмотический градиент между канальцевой и интерстициальной жидкостью, служащий мощной движущей силой для обратного входа в каналец NaCl (или выхода из него воды). Это может свести на нет и без того незначительную гиперосмолярность, создаваемую в интерстиции.
Итак, реабсорбция осмотически активных веществ из канальца в интерстиций без дополнительных механизмов была бы крайне неэффективным способом концентрирования интерстиция.
Для повышения концентрирования интерстиция петля Генле работает как поворотно-противоточный множитель. Рассмотрим сначала упрощенную схему его работы.
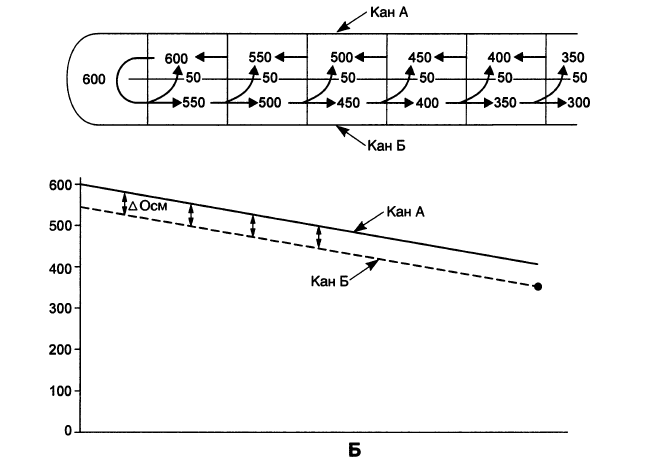
На рис. 15.8, Б приведена типичная поворотно-противоточная система, имеющая вид U-образной петли. Жидкость поступает в каналец А, движется до изгиба петли, заворачивает (отсюда — поворотная) и течет в канальце Б в противоположном направлении (отсюда — противоточная). Вновь разделим нашу систему на условные квадратики; в каждом из них из канальца Б в каналец А поступает 50 мосмоль/л NaCl. Поскольку жидкость в канальце А движется, то, переходя из квадратика в квадратик, она все больше обогащается NaCl, то есть становится все гипертоничнее. Совершив поворот, она поступает в каналец Б, где происходит активное выкачивание NaCl; двигаясь по этому канальцу, жидкость в каждом квадратике теряет 50 мосмоль/л NaCl и тем самым становится все гипотоничнее.
В результате, как видно из графика на рис. 15.8, Б: к изгибу петли достигается высокая осмолярность жидкости в обоих канальцах, и чем длиннее будет петля, тем большая осмолярность будет достигнута; осмотический градиент между канальцами постоянен (одинаков на уровне каждого квадратика) и невелик.
Реальная же поворотно-противоточная система почек отличается следующими особенностями.
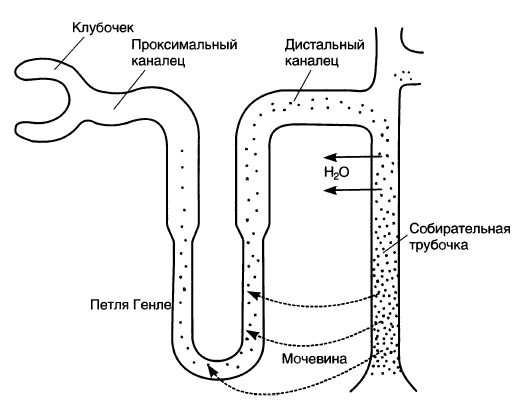
1. В толстом сегменте восходящей части петли Генле активно реабсорбируются электролиты (Nа+, К+ и Сl-), но он не проницаем для воды. Напротив, тонкий нисходящий сегмент непроницаем для электролитов, но высоко проницаем для воды; поэтому электролиты не входят в него, но вместо этого им навстречу по градиенту осмотического давления выходит вода. Понятно, что с точки зрения повышения осмолярности вход электролитов и выход воды равноценны.
2. Обмен между восходящей и нисходящей частями петли Генле осуществляется не непосредственно, а через интерстиций. Поскольку нисходящая часть петли Генле высоко проницаема для воды, осмолярность в интерстиции становится такой же, как в этой части, то есть постепенно повышается к изгибу петли. В этом концентрировании интерстиция, как уже говорилось, и заключается цель работы петли Генле.
2. Реабсорбируемые в восходящей части электролиты и выходящая из нисходящей части вода поступают в капилляры (прямые сосуды) и относятся с током крови. Поэтому петля Генле не только создает высокую осмолярность интерстиция, но также способствует дальнейшей реабсорбции воды и электролитов: из 60 л/сут жидкости, поступающей из проксимального канальца, 40 л/сут реабсорбируются в петле Генле.
3. Рассмотренный нами механизм концентрирования интерстиция, основанный на активной реабсорбции Nа+ и других электролитов, работает в верхних отделах петли Генле — там, где восходящая часть представлена толстым сегментом. В тонком сегменте (нисходящем и восходящем) процессы активного транспорта не идут; следовательно, данный механизм здесь невозможен. Тем не менее и здесь осмолярность интерстиция продолжает нарастать, но обусловлено это иным — мочевинным — механизмом концентрирования мочи. Суть этого механизма в следующем:
1. Поздние отделы дистальных извитых канальцев и корковые отделы собирательных трубочек в присутствии АДГ проницаемы для воды.
2. Поскольку жидкость в них гипотонична, а тоничность окружающего интерстиция по направлению к глубинным слоям постепенно нарастает, вода выходит в интерстиций. Эти отделы непроницаемы для мочевины; по мере выхода воды концентрация мочевины в канальцевой жидкости нарастает и в мозговом отделе собирательных трубочек становится очень высокой;
глубинные отделы собирательных трубочек проницаемы для мочевины; мочевина выходит в интерстиций по концентрационному градиенту, увеличивая тем самым его осмолярность;
3. Из интерстиция мочевина частично входит обратно в канальцы в области тонкого сегмента, так как: этот сегмент для нее проницаем; ее концентрация в интерстиции мозгового вещества очень высока. Тем самым мочевина совершает кругооборот, благодаря которому, с одной стороны, постоянно поддерживается высокая осмолярность мозгового вещества, с другой — большая часть мочевины выделяется с мочой.
В результате совместной работы натриевого и мочевинного механизмов концентрирования мочи в глубинных слоях интерстиция достигается осмолярность порядка 1200 мосмоль/л. Если собирательная трубочка, проходящая через эти слои, будет проницаема для воды, то вода выйдет в интерстиций до выравнивания осмотического градиента, и осмолярность мочи достигнет максимального значения — 1200мосмоль/л. Именно это и происходит под действием АДГ.
Действие АДГ
Вырабатывается в гипоталамусе; хранится в нейрогипофизе; выбрасывается в ответ на стимуляцию осморецепторов гипоталамуса (то есть на повышение осмолярности крови), а также в ответ на существенное снижение импульсации от волюмо- и барорецепторов (то есть на снижение объема крови и артериального давления); действуя на почки, повышает реабсорбцию воды и, следовательно, по механизму отрицательной обратной связи снижает осмолярность крови и повышает ее объем.
Механизм действия АДГ на почки заключается в том, что этот гормон повышает проницаемость для воды поздних отделов дистальных извитых канальцев и собирательных трубочек:
в поздних отделах дистальных извитых канальцев под действием АДГ вода начинает выходить вслед за активно реабсорбируемым Nа+, и в результате уменьшается степень разбавления мочи;
в собирательных трубочках под действием АДГ вода начинает выходить в гипертоничный интерстиций, достигая в конечном счете предельной осмолярности — 1200 мосмоль/л;
кроме того, в присутствии АДГ срабатывает мочевинный механизм концентрирования мочи.
Резюме: механизмы концентрирования и разбавления мочи
1. Петля Генле, работая как поворотно-противоточная система, создает высокую гипертоничность интерстиция.
2.В собирательную трубочку поступает гипотоничная жидкость.
3. Собирательная трубочка проходит через интерстиций, который по направлению к глубинным отделам становится все более гипертоничным; если собирательная трубочка не проницаема для воды, из нее будет оттекать та же гипотоничная жидкость, что в нее поступает; если она проницаема для воды, то вода будет реабсорбироваться, а оттекать будет гипертоничная моча.
4. Гормоном, повышающим проницаемость собирательной трубочки для воды, является АДГ.
Регуляция выведения воды при разных водных нагрузках
1. При максимальной водной нагрузке (например, потреблении 12 л чистой воды в сутки): кровь становится гипоосмолярной; уменьшается стимуляция осморецепторов гипоталамуса; прекращается выделение АДГ; в отсутствие АДГ вода в собирательных трубочках не реабсорбируется и выделяется большой объем (12 л/сут) резко гипотоничной (50 мосмоль/л) мочи.
Это состояние называется водным диурезом.
2. При минимальной водной нагрузке (отсутствие приема воды, сухоядение): кровь становится гиперосмолярной; стимулируются осморецепторы гипоталамуса; выделение АДГ становится максимальным; при максимальной секреции АДГ вода в собирательных трубочках реабсорбируется до достижения осмотического равновесия с интерстицием, и выделяется малый объем (0,5 л/сут) резко гипертоничной мочи (1200 мосмоль/л).
Это состояние называется антидиурезом.