

Билет 1
1. По своему строению митохондрии представляют собой органеллы, обычно сферической формы, встречающиеся в эукариотической клетке в количестве от нескольких сотен до 1—2 тысяч и занимающие 10—20% её внутреннего объёма. Митохондрии способны перемещаться по цитоплазме в зоны наибольшего энергопотребления.
На наружную мембрану приходится около 7% площади поверхности всех мембран клеточных органелл. Основная функция наружной мембраны — отграничение митохондрии от цитоплазмы. Наружная мембрана митохондрии состоит из липидов с вкраплениями белков. Особую роль играет порин — каналообразующий белок. Он образует в наружной мембране отверстия диаметром 2-3 нм, через которые могут проникать небольшие молекулы и ионы весом до 5 кДа. Для наружной мембраны характерно присутствие ферментов: монооксигеназы, ацил-СоА-синтетазы и фосфолипазы А2. Наружная мембрана митохондрии может взаимодействовать с мембраной эндоплазматического ретикулума; это играет важную роль в транспортировке липидов и ионов кальция.
Внутренняя мембрана состоит в основном из белковых комплексов (соотношение белок/липид — 3:1) и образует многочисленные гребневидные складки — кристы, существенно увеличивающие площадь её поверхности и, например, в клетках печени составляет около трети всех клеточных мембран. Характерной чертой состава внутренней мембраны митохондрий является присутствие в ней кардиолипина — особого фосфолипида, содержащего сразу четыре жирные кислоты и делающего мембрану абсолютно непроницаемой для протонов
Локализации ферментов митохондрий
1). Наружная мембрана содержит: а) элонгазы, ферменты удлиняющие молекулы насыщенных жирных кислот; б) кинуренингидроксилазу; в) моноаминооксидазу (маркер) и др.
2). Межмембранное пространство содержит: а) аденилатциклазу; б) нуклеозиддифосфаткиназы.
3). Внутренняя мембрана содержит: а) ферменты цепи окислительного фосфорилирования, из них цитохромоксидаза - маркер; б) СДГ в) β-оксибутират ДГ; г). карнитинацилтрансферазу.
4). Матрикс содержит: а) ферменты ЦТК; б) ферменты β-окисления жирных кислот; в) аминотрансферазы АСТ, АЛТ; г) глутамат ДГ д) фосфоенолпируваткарбоксилазу е). пируват ДГ
Все ферменты митохондриального окисления встроены во внутреннюю мембрану митохондрий. Только первый переносчик протонов и электронов – никотинамидная дегидрогеназа расположена в матриксе митохондрии. Этот фермент отнимает водород от субстрата и передает его следующему переносчику. Полный комплекс таких ферментов образует “дыхательный ансамбль” («дыхательную цепь»), в пределах которого атомы водорода отнимаются от субстрата, затем передаются последовательно от одного переносчика к другому, и, наконец, передаются на кислород воздуха с образованием воды. Существует строгая последовательность работы каждого звена в цепочке переносчиков. Эта последовательность определяется величиной редокс-потенциала. Вещества с положительным ОВП окисляют водород (отнимают от него электроны), вещества с отрицательным ОВП окисляются самим водородом.
2.
![]() -
суммарное
уравнение, отражающее окислительное
декарбоксилирование пирувата,
восстановление НАД до НАДН и образование
ацетил-SKoА.
-
суммарное
уравнение, отражающее окислительное
декарбоксилирование пирувата,
восстановление НАД до НАДН и образование
ацетил-SKoА.
 Суть
первых трех реакций сводится к
декарбоксилированию пирувата
(катализируется пируватдегидрогеназой,
Е1),
окислению полученного гидроксиэтила
до ацетила и переносу ацетила на коэнзим
А (катализируется дигидролипоат-ацетилтрансферазой,
Е2).
Суть
первых трех реакций сводится к
декарбоксилированию пирувата
(катализируется пируватдегидрогеназой,
Е1),
окислению полученного гидроксиэтила
до ацетила и переносу ацетила на коэнзим
А (катализируется дигидролипоат-ацетилтрансферазой,
Е2).
 Оставшиеся
2 реакции необходимы для возвращения
липоевой кислоты и ФАД в окисленное
состояние (катализируются
дигидролипоат-дегидрогеназой, Е3).
При этом образуется НАДН.
Оставшиеся
2 реакции необходимы для возвращения
липоевой кислоты и ФАД в окисленное
состояние (катализируются
дигидролипоат-дегидрогеназой, Е3).
При этом образуется НАДН.
Биологическое значение: является важным этапом катаболизма, позволяющим включаться в цикл Кребса тем веществам, при распаде которых образуется ПВК. Образовавшаяся моелкула НАДН2, окисляется в длинной дыхательной цепи с образованием 3х молекул АТФ.
3. Общая характеристика: представляет собой магистральный, циклический, метаболический путь, в котором происходит окисление активной уксусной кислоты в некоторых других соединений образующихся при распаде углеводов, липидов, белков и который обеспечивает дыхательную цепь восстановительными коферментами.
Энергетическое значение: за один оборот цитратного цикла синтезируется 12 молекул АТФ. Девять из низ образуется за счет энергии транспорта в дыхательной цепи трех пап водорода от трех молекул НАДН+Н+. Две молекулы АТР, синтезируются при окислении 1 молекулы FADH2, так как в дыхательной цепи в данном случае действует только два пункта сопряжения с окислительным фосфорилированием ADP.
Билет 2
1. Метаболизм: происходит в клетках и тканях и заключается в осуществлении химических превращений, ведущих к синтезу и распаду химических соединений, что сопровождается образованием промежуточных и конечных продуктов обмена. Анаболизм и катаболизм.
Процессы анаболизма и катаболизма взаимосвязаны. Например, вещества, образующиеся в ходе катаболизма (например, глюкоза, глицерин, жирные кислоты, аминокислоты) могут использоваться организмом для биосинтетических процессов. Это явление получило название амфиболизма
Биологическое значение метаболизма состоит в том что бы: снабдить клетку энергией, обеспечить строительными блоками белков, собрать макромолекулы для построения клеточных структур и обеспечить распад функционально активных молекул.
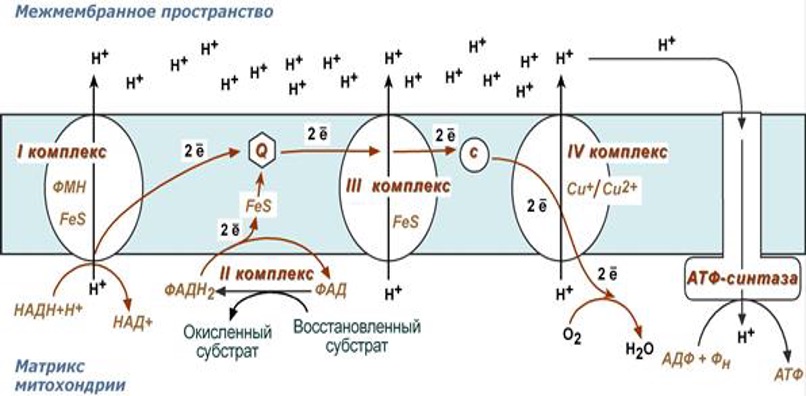
2. Дыхательная цепь (ферменты тканевого дыхания) - это переносчики протонов и электронов от окисляемого субстрата на кислород.
Дыхательная цепь состоит из:
1. НАД - зависимой дегидрогеназы;
2. ФАД- зависимой дегидрогеназы;
3. Убихинона (КоQ);
4. Цитохрмов b, c, a+a3 .
5. белков, содержащих негиминовое железо.
2 комплекс. ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует, его выделение условно. К нему относятся ФАД-зависимые ферменты, расположенные на внутренней мембране – например, ацил-SКоА-дегидрогеназа (β-окисление жирных кислот), сукцинатдегидрогеназа (цикл трикарбоновых кислот), митохондриальная глицерол-3-фосфат-дегидрогеназа (челночный механизм переноса атомов водорода).
Функция
Восстановление ФАД в окислительно-восстановительных реакциях.
Обеспечение передачи электронов от ФАДН2 на железосерные белки внутренней мембраны митохондрий. Далее эти электроны попадают на коэнзим Q (убихинон).
3 комплекс. КоQ-цитохром c-оксидоредуктаза
По другому данный комплекс называется цитохром с редуктаза. В его составе имеются молекулы цитохрома b и цитохрома c1, железо-серные белки. Комплекс представляет собой 2 мономера, в каждом из которых насчитывается 11 полипептидных цепей.
Функция
Принимает электроны от коэнзима Q и передает их на цитохром с.
Переносит 2 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
Имеются разногласия по поводу количества переносимых ионов H+ при участии 3-го и 4-го комплексов. По одним данным, третий комплекс переносит 2 иона H+ и четвертый комплекс переносит 4 иона H+. По другим авторам, наоборот, третий комплекс переносит 4 иона H+ и четвертый комплекс переносит 2 иона H+.
4 комплекс. Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а и а3, он называется также цитохромоксидаза, состоит из 13 субъединиц. В комплексе имеются ионы меди, соединенные с белками комплекса через HS-группы цистеина, и формирующие центры, подобные тем, что имеются в железо-серных белках.
Функция
Принимает электроны от цитохрома с и передает их на кислород с образованием воды.
Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
3.
 В
реакции образования цитрата углеродный
атом метильной труппы ацетил-КоА
связывается с карбонильной группой
оксалоацетата; одновременно расщепляется
тиоэ-фирная связь и освобождается
коэнзим A. Катализирует реакцию цитрат
синтаза, фермент, локализованный в
матриксе митохондрий. Цитратсинтаза
локализуется в матриксе митохондрий,
ее активируют ЩУК, НАД+; ингибируют АТФ,
НАДН2, Сукцинил-КоА, цитрат.
В
реакции образования цитрата углеродный
атом метильной труппы ацетил-КоА
связывается с карбонильной группой
оксалоацетата; одновременно расщепляется
тиоэ-фирная связь и освобождается
коэнзим A. Катализирует реакцию цитрат
синтаза, фермент, локализованный в
матриксе митохондрий. Цитратсинтаза
локализуется в матриксе митохондрий,
ее активируют ЩУК, НАД+; ингибируют АТФ,
НАДН2, Сукцинил-КоА, цитрат.
В реакции образования цитрата углеродный атом метильной группы ацетил-КоА связывается с карбонильной группой оксалоацетата; одновременно расщепляется тио-эфирная связь и освобождается коэнзим А.
4) Витамин В2, рибофлавин, витамин роста. Он включает флавин и рибитол, распространён в оболочке злаков, в дрожжах. Суточная потребность 1-2 мг. Строение и роль коферментов.
входит в состав коферментов ФМН и ФАД, участвует в биологическом окислении. Авитаминоз проявляется в виде дерматита, катаракты, анемии, поражении сердечной мышцы
Билет 3
1)Обмен веществ или метаболизм – это совокупность химических реакций в организме, которые обеспечивают его веществами и энергией, необходимыми для жизнедеятельности. (совокупность химических процессов, которым подвергаются соединения с момента их поступления в организм и до выделения конечных продуктов обмена.) Биологическое значение: Энергетическая – снабжение клетки химической энергией, Пластическая – синтез макромолекул как строительных блоков, Специфическая – синтез и распад биомолекул, необходимых для выполнения специфических клеточных функций
2) Принцип работы дыхательной цепи 1. Образующиеся в реакциях катаболизма НАДН и ФАДН2 передают атомы водорода (т.е. протоны водорода и электроны) на ферменты дыхательной цепи. 2. Электроны движутся по ферментам дыхательной цепи и теряют энергию. 3. Эта энергия используется на выкачивание протонов Н+ из матрикса в межмембранное пространство. 4. В конце дыхательной цепи электроны попадают на кислород и восстанавливают его до воды. 5. Протоны Н+ стремятся обратно в матрикс и проходят через АТФ-синтазу. 6. При этом они теряют энергию, которая используется для синтеза АТФ.
3) Цитратсинтаза локализуется в матриксе митохондрий, ее активируют ЩУК, НАД+; ингибируют АТФ, НАДН2, Сукцинил-КоА, цитрат.

В реакции образования цитрата углеродный атом метильной группы ацетил-КоА связывается с карбонильной группой оксалоацетата; одновременно расщепляется тио-эфирная связь и освобождается коэнзим А.
Билет 4
1) Весь катаболизм условно подразделяется на три этапа, включающие реакции общих и специфических путей.
Первый этап Происходит в кишечнике (переваривание пищи) или в лизосомах (самообновление клеток) при расщеплении уже ненужных или лишних молекул. При этом освобождается около 1% энергии, заключенной в молекуле. Она рассеивается в виде тепла.
Второй этап Вещества, образованные при внутриклеточном гидролизе или проникающие в клетку из крови, на втором этапе обычно превращаются -в пировиноградную кислоту (моносахариды в гликолизе), -в ацетил-SKoA, в пируват и другие кетокислоты (в катаболизме аминокислот), -в ацетил-SКоА (при β-окислении жирных кислот).
Локализация второго этапа – цитозоль и митохондрии. Часть полученной энергии рассеивается в виде тепла и примерно 13% энергии вещества усваивается, т.е. запасается в виде макроэргических связей АТФ.
Третий этап Все реакции этого этапа идут в митохондриях. Ацетил-SКоА (и кетокислоты) включается в реакции цикла трикарбоновых кислот, где углероды веществ окисляются до углекислого газа.
Выделенные атомы водорода соединяются с НАД и ФАД и восстанавливают их. После этого НАДН и ФАДН2 переносят водород в цепь ферментов дыхательной цепи, расположенную на внутренней мембране митохондрий. Здесь в результате процесса под названием "окислительное фосфорилирование" образуется вода и главный продукт биологического окисления – АТФ.
Часть выделенной на этом этапе энергии молекулы рассеивается в виде тепла и около 46% энергии исходного вещества усваивается, т.е. запасается в связях АТФ и ГТФ.
2) окислительное декарбоксилирование а-кетоглутарата 3)3 комплекс. КоQ-цитохром с-оксидоредуктаза
По другому данный комплекс называется цитохром с редуктаза. В его составе имеются молекулы цитохрома b и цитохрома c1, железо-серные белки. Комплекс представляет собой 2 мономера, в каждом из которых насчитывается 11 полипептидных цепей.
Функция Принимает электроны от коэнзима Q и передает их на цитохром с. Переносит 2 иона Н+ на наружную поверхность внутренней митохондриальной мембраны. !!!!Имеются разногласия по поводу количества переносимых ионов H+ при участии 3-го и 4-го комплексов. По одним данным, третий комплекс переносит 2 иона H+ и четвертый комплекс переносит 4 иона H+. По другим авторам, наоборот, третий комплекс переносит 4 иона H+ и четвертый комплекс переносит 2 иона H+.
4 комплекс. Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а и а3, он называется также цитохромоксидаза, состоит из 13 субъединиц. В комплексе имеются ионы меди, соединенные с белками комплекса через HS-группы цистеина, и формирующие центры, подобные тем, что имеются в железо-серных белках.
Функция Принимает электроны от цитохрома с и передает их на кислород с образованием воды. Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
Билет №5
1) БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ (БО) - это совокупность окислительных процессов в живом организме, протекающих с обязательным участием кислорода. Синоним - ТКАНЕВОЕ ДЫХАНИЕ. Окисление одного вещества невозможно без восстановления другого вещества. Окислительно-восстановительных процессов в живой природе очень много. Часть окислительно-восстановительных процессов, протекающих с участием кислорода, относится к биологическому окислению.
Функции биологического окисления
· Энергетическое обеспечение: поддержания температуры тела; биолюминесценции (свечения); химических синтезов; осмотических явлений; электрических процессов; механической работы.
· Синтез важнейших (ключевых) метаболитов.
· Регуляция обмена веществ.
· Устранение вредных для клетки продуктов обмена (шлаков).
· Детоксикация проникших в организм чуждых соединений – ксенобиотиков (пестицидов, препаратов бытовой химии, лекарственных средств, промышленных загрязнений и т.п.).
Типы биологического окисления
Существуют 2 типа биологического окисления:
Свободное окисление - Окисление, не сопряженное с фосфорилированием АДФ и не сопровождающееся трансформацией энергии, выделяющейся при окислении, в энергию макроэргических связей. При свободном окислении высвобождающаяся энергия переходит в тепловую и рассеивается.
Окисление, сопряженное с фосфорилированием АДФ. Этот тип биологического окисления осуществляется двумя путями: субстратное фосфорилирование, окислительное фосфорилирование
2) · Комплекс II окисляет сукцинат до фумарата и восстанавливает убихинон:
Сукцинат + Q → Фумарат + QH2
· Цитохро́м-bс1-ко́мплекс (комплекс цитохромов bc1) или убихинол-цитохром с-оксидоредуктаза, или комплекс III — мультибелковый комплекс дыхательной цепи переноса электронов и важнейший биохимический генератор протонного градиента на мембране митохондрий. Этот мультибелковый трансмембранный комплекс кодируется митохондриальным (цитохром b) и ядерным геномами.
· Цитохром с оксида́за (цитохромоксидаза) или цитохром с-кислород-оксидоредуктаза, также известная как цитохром aa3 и комплекс IV — терминальная оксидаза аэробной дыхательной цепи переноса электронов, которая катализирует перенос электронов с цитохрома с на кислород с образованием воды. Цитохромоксидаза присутствует во
внутренней мембране митохондрий всех эукариот, где её принято называть комплекс IV, а также в клеточной мембране многих аэробных бактерий.
3) В результате окисления сукцината до оксалоацетата будет образовываться 5 молекул АТФ.
Сукцинат (янтарная кислота) в ходе нескольких реакций (окисление, гидратация, окисление) превращается в оксалоацеат. Данная последовательность превращений представляет собой процесс b-окисления. Реакции последней стадии ЦТК представляют собой регенерирование оксалоацетата. Окисление сукцината катализирует сукцинатдегидрогеназа. Этот фермент непосредственно связан с цепью переноса электронов, и в отличие от других ферментов ЦТК является интегральным белком внутренней мембраны митохондрий.
Продукт окисления сукцината — фумарат — гидратируется фумаразой. Происходит стереоспецифическое присоединение водорода и гидроксила с образованием L-малата (яблочной кислоты). Наконец, L-малат окисляется в оксалоацетат малатдегидрогеназой.
Билет №6
1) Весь катаболизм условно подразделяется на три этапа, включающие реакции общих и специфических путей.
Первый этап Происходит в кишечнике (переваривание пищи) или в лизосомах (самообновление клеток) при расщеплении уже ненужных или лишних молекул. При этом освобождается около 1% энергии, заключенной в молекуле. Она рассеивается в виде тепла.
Второй этап Вещества, образованные при внутриклеточном гидролизе или проникающие в клетку из крови, на втором этапе обычно превращаются -в пировиноградную кислоту (моносахариды в гликолизе), -в ацетил-SKoA, в пируват и другие кетокислоты (в катаболизме аминокислот), -в ацетил-SКоА (при β-окислении жирных кислот).
Локализация второго этапа – цитозоль и митохондрии. Часть полученной энергии рассеивается в виде тепла и примерно 13% энергии вещества усваивается, т.е. запасается в виде макроэргических связей АТФ.
Третий этап Все реакции этого этапа идут в митохондриях. Ацетил-SКоА (и кетокислоты) включается в реакции цикла трикарбоновых кислот, где углероды веществ окисляются до углекислого газа.
Выделенные атомы водорода соединяются с НАД и ФАД и восстанавливают их. После этого НАДН и ФАДН2 переносят водород в цепь ферментов дыхательной цепи, расположенную на
внутренней мембране митохондрий. Здесь в результате процесса под названием "окислительное фосфорилирование" образуется вода и главный продукт биологического окисления – АТФ.
Часть выделенной на этом этапе энергии молекулы рассеивается в виде тепла и около 46% энергии исходного вещества усваивается, т.е. запасается в связях АТФ и ГТФ.
2) Цитохромы в дыхательной цепи выстроены в порядке возрастания окислительновосстановительного потенциала. Они представляют собой гемопротеины, в которых простетическая геминовая группа близка к гему гемоглобина (у цитохрома b идентична). Ионы железа в составе гема при получении и отдаче электронов обратимо изменяют свою валентность.
В процессах тканевого дыхания наиболее важную роль играют цитохромы b, с , с, а и а . Цитохром а3 представляет собой терминальный участок 1 3 3 дыхательной цепи – цитохромоксидазу, которая осуществляет окисление цитохрома с и образование воды. Элементарный акт представляет собой двухэлектронное восстановление одного атома кислорода, т.е. каждая молекула кислорода одновременно взаимодействует с двумя электрон-транспортными цепями. При транспорте каждой пары электронов во внутримитохондриальном пространстве может накапливаться до 6 протонов. Строение дыхательной цепи интенсивно исследуется. В числе последних достижений молекулярной биохимии – установление тонкой структуры дыхательных ферментов с помощью рентгеноструктурного анализа. С помощью электронного микроскопа с наивысшим доступным в настоящее время разрешением можно «увидеть» структуру цитохромоксидазы.
KoQ (убихинон), необходимый компонент дыхательной цепи, является производным бензохинона с боковой цепью, которая у млекопитающих чаще всего представлена 10 изопреноидными единицами. Как любой хинон, KoQ способен
находиться и в восстановленном, и окисленном состоянии. Это свойство определяет его роль в дыхательной цепи - служить коллектором восстановительных эквивалентов, поставляемых в дыхательную цепь через флавиновые дегидрогеназы. Содержание его значительно превосходит содержание других компонентов дыхательной цепи.
3) В результате окисления a-кетоглутарата и сукцината до оксалоацетата будет образовываться 9 молекул АТФ.
На третьей стадии a-кетоглутарат также подвергается окислительному декарбоксилированию. Эту реакцию катализирует организованный ансамбль ферментов — a-кетоглутарат-дегидрогеназный комплекс. Состав ансамбля сходен с комплексом, катализирующим окислительное декарбоксилирование пирувата, и оба процесса имеют множество общих деталей. Продуктом реакции является четырехуглеродное активированное соединение —сукцинил-СоА. Макроэргическая связь сукцинильного тиоэфира расщепляется с участием сукцинаттиокиназы, и это сопряжено с фосфорилированием GDP (у млекопитающих) или АТР (у бактерий, высших растений)
Образовавшийся на предыдущей стадии сукцинат (янтарная кислота) в ходе нескольких реакций (окисление, гидратация, окисление) превращается в оксалоацеат. Данная последовательность превращений представляет собой процесс b-окисления. Реакции последней стадии ЦТК представляют собой регенерирование оксалоацетата. Окисление сукцината катализирует сукцинатдегидрогеназа. Этот фермент непосредственно связан с цепью переноса электронов, и в отличие от других ферментов ЦТК является интегральным белком внутренней мембраны митохондрий.
Продукт окисления сукцината — фумарат — гидратируется фумаразой. Происходит стереоспецифическое присоединение водорода и гидроксила с образованием L-малата (яблочной кислоты). Наконец, L-малат окисляется в оксалоацетат малатдегидрогеназой.
Билет №7
1. Питание - это основная биологическая потребность любого живого организма. Оттого насколько правильно питается человек зависит его жизнеспособность,‘работоспособность, устойчивость организма к воздействиям вредных факторов. Питание обеспечивает энергетический обмен веществ - снабжение организма питательными веществами, чтобы восстанавливать те вещества, которые были израсходованы организмом в процессе жизнедеятельности. Питание обеспечивает пластический процесс -восстановление израсходованных и синтез новых клеточных элементов, а также откладывание веществ в тканях: жир откладывается в
жировой клетчатке; гликоген - в печени. Энергетические расходы организма не требуют
какого-либо качественного состава пищи, поскольку отдельные пищевые элементы
(жиры, белки, углеводы) могут быть заменены по принципу их изодинамического
равновесия, те., по их калорийности. Но, для пластических процессов необходимо
наличие определенного минимума определенных компонентов пищи, те. важен
качественный состав пищи. Два основных процесса, руководящие обменом веществ в
организме - это ассимиляция и диссимиляция, которые протекают параллельно.
2.
Высокоэнергетические фосфаты- группа органических фосфатов, гидролиз которых приводит к освобождению большого количества свободной энергии
К группе высокоэнергетических фосфатов, помимо АТФ, относят енолфосфаты, ангидриды и фосфогуанидины.
АТФ - молекула, богатая энергией, поскольку она содержит две фосфоаншдридные связи. При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и ортофосфат Рi При этом изменение свободной энергии составляет -7,3 ккал/моль. При условиях, существующих в клетке в норме (рН 7,0, температура 37 °С), фактическое значение ΔG0' для процесса гидролиза составляет около -12 ккал/моль. Величина свободной энергии гидролиза АТФ делает возможным его образование из АДФ за счёт переноса фосфатного остатка от таких высокоэнергетических фосфатов, как, например, фосфоенолпируват 1или 1,3-бисфосфоглицерат; в свою очередь, АТФ может участвовать в таких эндергонических реакциях, как фосфорилирование глюкозы или глицерина. АТФ выступает в роли донора энергии в эндергонических реакциях многих анаболических процессов. Некоторые биосинтетические реакции в организме могут протекать при участии других нуклеозидтрифосфатов, аналогов АТФ; к ним относят гуанозинтрифосфат (ГТФ), уридинтрифосфат (УТФ) и цитидинтрифосфат (ЦТФ). Все эти нуклеотиды, в свою очередь, образуются при использовании свободной энергии концевой фосфатной группы АТФ. Наконец, за счёт свободной энергии АТФ совершаются различные виды работы, лежащие в основе жизнедеятельности организма, например, такие как мышечное сокращение или активный транспорт веществ.
Таким образом, АТФ - главный, непосредственно используемый донор свободной энергии в биологических системах. В клетке молекула АТФ расходуется в течение одной минуты после её образования. У человека количество АТФ, равное массе тела, образуется и разрушается каждые 24 ч.
Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счёт энергии окисления органических
соединений. Цикл АТФ-АДФ - основной механизм обмена энергии в биологических системах, а АТФ - универсальная "энергетическая валюта".
3. Известны два субстрата. Главным из них является сукцинат (янтарная кислота).
Сукцинатдегидрогеназа – это комплекс II, который в укороченном варианте цепи является начальным звеном окисления. В составе комплекса – простетическая группа ФАД и FeSII. От ФАД.Н2 два атома водорода переносятся на KoQ. Значит, первое звено, которое имеется в полной цепи - исключается. Перепад окислительно-восстановительного потенциала между ФАД и KoQ невелик. Поэтому переноса H+ в межмембранное пространство в этой точке не происходит. DmH+ также создается, но меньший, чем в полной цепи. Значит, меньше и эффективность фосфорилирования - коэффициент Р/О=2.
Аналогичным образом окисляется и второй субстрат – ацил-КоА (активная форма любой жирной кислоты)
Билет 8
1. Образование углекислого газа в организме.
СО2 в организме образуется двумя путями - путем прямого и окислительного декарбоксилирования. Основная масса СО2 образуется в ЦТК. Первая молекула СО2 образуется путем прямого декарбоксилирования изоцитрата. Вторая молекула СО2 путем окислительного декарбоксилирования альфа-кетоглутората. Одна молекула СО2 образуется в результате окислительного декарбоксилирования ПВК. Человек за сутки выделяет около 500 мл СО2.
В ЦТК есть только два типа декарбоксилирования - прямое и окислительное. 1. Прямое декарбоксилирование изоцитрата под действием изоцитратдегидрогеназы, которая обладает декарбоксилирующим эффектом, превращается в альфа-кетоглуторат и НАДН, который является источником водорода для дыхательной цепи, окисление которого приводит к образованию 3х АТФ и воды. Кроме того в этой реакции образуется СО2. 2. Окислительное декарбоксилирование альфа-кетоглутората происходит под действием альфа-кетоглуторатдегидрогеназного комплекса, который включает три фермента и пять кофакторов - ТДФНSКоА, НАД, ФАД, липоевая кислота. Продуктом реакции является образование макроэргического соединения - сукцинил-КоА. В результате этой реакции образуется еще одна молекула восстановительного эквивалента НАДН и СО2.
2. Характеристика и роль цитохромов в дыхательной цепи.
3 комплекс. КоQ-цитохром с-оксидоредуктаза
По-другому данный комплекс называется цитохром с редуктаза. В его составе имеются молекулы цитохрома b и цитохрома c1, железо-серные белки. Комплекс представляет собой 2 мономера, в каждом из которых насчитывается 11 полипептидных цепей.
Функция:
1. Принимает электроны от коэнзима Q и передает их на цитохром с.
2. Переносит 2 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
Имеются разногласия по поводу количества переносимых ионов H+ при участии 3-го и 4-го комплексов. По одним данным, третий комплекс переносит 2 иона H+ и четвертый комплекс переносит 4 иона H+. По другим авторам, наоборот, третий комплекс переносит 4 иона H+ и четвертый комплекс переносит 2 иона H+.
4 комплекс. Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а и а3, он называется также цитохромоксидаза, состоит из 13 субъединиц. В комплексе имеются ионы меди, соединенные с белками комплекса через HS-группы цистеина, и формирующие центры, подобные тем, что имеются в железо-серных белках.
Функция:
1. Принимает электроны от цитохрома с и передает их на кислород с образованием воды.
2. Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
3. Тканевое дыхание — процесс окисления водорода кислородом до воды ферментами дыхательной цепи. Регуляция тканевого дыхания происходит в 3 этапа: 1) окислительное образование ацетилкофермента А из пировиноградной кислоты, жирных кислот и аминокислот 2) разрушение ацетильных остатков в трикарбоновых кислот цикле с освобождением 2 молекул углекислого газа и 4 пар атомов водорода, частично акцептируемых коферментами никотинамидадениндинуклеотидом и флавинадениндинуклеотидом и частично переходящих в раствор в виде протонов 3) перенос электронов и протонов к молекулярному кислороду (образование H2O) — процесс, катализируемый набором дыхательных ферментов и сопряжённый с образованием АТФ (энергии). Первые две стадии подготавливают третью, в ходе которой в результате последовательных окислительно-восстановительных реакций происходит освобождение основной части энергии, вырабатываемой в клетке. При этом около 50% энергии в результате окислительного фосфорилирования запасается в форме богатых энергией связей АТФ, а остальная часть её выделяется в виде тепла. Тканевое дыхание обеспечивает образование и постоянное пополнение АТФ в клетках.
Нарушение тканевого дыхания связано с повреждением внутренней мембраны митохондрий, химическими и физическими факторами приводит разобщению окислительного фосфорилирования, нарушению синтеза АТФ, торможению анаболических реакций, межмембранного транспорта и всех видов обмена веществ.
Дыхательный контроль.
Дыхательный контроль – это прямое влияние электрохимического градиента на скорость движения электронов по дыхательной цепи (т.е. на величину дыхания). В свою очередь, величина градиента напрямую зависит от соотношения АТФ / АДФ, количественная сумма которых в клетке практически постоянна ([АТФ] + [АДФ] = const). Реакции катаболизма направлены на поддержание постоянно высокого уровня АТФ и низкого АДФ.
Возрастание протонного градиента возникает при снижении количества АДФ и накоплении АТФ (состояние покоя), т.е. когда АТФ-синтаза лишена своего субстрата и ионы Н+ не проникают в матрикс митохондрии. При этом ингибирующее влияние градиента усиливается и продвижение электронов по цепи замедляется. Ферментные комплексы остаются в восстановленном состоянии. Следствием является уменьшение окисления НАДН и ФАДН2 на I и II комплексах, ингибирование ферментов ЦТК при участии НАДН и замедление катаболизма в клетке. Снижение протонного градиента возникает при исчерпании резервов АТФ и избытке АДФ, т.е. при работе клетки. В этом случае активно работает АТФ-синтаза и через канал Fо проходят в матрикс ионы Н+. При этом протонный градиент, естественно, снижается, поток электронов по цепи возрастает, и в результате повышается выкачивание ионов Н+ в межмембранное пространство и снова их быстрое "проваливание" через АТФ-синтазу внутрь митохондрий с синтезом АТФ. Ферментные комплексы I и II усиливают окисление НАДН и ФАДН2 (как источников электронов) и снимается ингибирующее влияние НАДН на цикл лимонной кислоты и пируватдегидрогеназный комплекс. Как итог – активируются реакции катаболизма углеводов и жиров
Разобщение дыхания и фосфорилирования.
Разобщение дыхания и фосфорилирования называют явление исчезновения на мембране электрохимического потенциала под действием разобщителей и прекращение синтеза АТФ. Разобщителями являются вещества, которые могут переносить протоны (протонофоры) или другие ионы (ионофоры) через мембрану минуя каналы АТФ-синтетазы. В результате разобщения количество АТФ снижается, АДФ увеличивается, возрастает скорость потребления О2, окисления НАДН2, ФАДН2, а образовавшаяся свободная энергия выделяется в виде теплоты. Как правило, разобщители — липофильные вещества, легко проходящие через мембраны. Например, вещество 2,4-динитрофенол (переносит Н+), лекарство - дикумарол, метаболит - билирубин, гормон щитовидной железы - тироксин, антибиотики - валиномицин и грамицидин
Билет 9
1.Самое важное макроэргическое соединение — АТФ. Используя энергию, заключенную в макроэргических связях АТФ. Образуется АТФ в процессах биологического окисления и при фотосинтезе. Энергия макроэргических связей используется для совершения любой работы: активации соединений,
синтеза биополимеров,
избирательного поглощения веществ из окружающей клетку среды и выброса из клетки ненужных продуктов,
мышечного сокращения и восстановления активного состояния организма.
Запас этих соединений позволяет организму быстро реагировать на изменение внешних условий и совершать физическую работу. При спортивной тренировке содержание макроэргических соединений в мышцах и скорость их образования возрастают.
2. В результате второй реакции образовавшаяся
лимонная кислота подвергается
дегидратированию с образованием
цис-аконитовой кислоты, которая,
присоединяя молекулу воды, переходит
в изолимонную кислоту (изоцитрат).
Катализирует эти обратимые реакции
гидратации– дегидратации фермент
аконитатгидратаза (аконитаза).
В результате второй реакции образовавшаяся
лимонная кислота подвергается
дегидратированию с образованием
цис-аконитовой кислоты, которая,
присоединяя молекулу воды, переходит
в изолимонную кислоту (изоцитрат).
Катализирует эти обратимые реакции
гидратации– дегидратации фермент
аконитатгидратаза (аконитаза).
3. Характеристика неполной (укороченной) дыхательной цепи.
Начинается от субстратов I типа: сукцината, ацил-КоА и α-глицерофосфата. Их дегидрирование происходит с помощью флавиновых ферментов, имеющих простетические группы ФАД и FeS центры. Ферменты являются интегральными белками внутренней мембраны митохондрий.
ФАД присоединяет два атома водорода от субстрата. Затем FeS-центры забирают 2ē, которые через КоQ поступают в цепь переноса электронов. Выталкивания протонов в межмембранное пространство не происходит, поскольку изменение свободной энергии незначительно.
Отличия полной цепи переноса электронов от укороченной: 1) субстраты полной ЦПЭ – II и III типов, укороченной – I типа; 2) в укороченной ЦПЭ отсутствуют пиридинзависимые дегидрогеназы.

Билет 10
1.Активные
формы кислорода химически очень
агрессивны: они повреждают белки и ДНК
и, главное, вызывают перекисное окисление
липидов - самоподдерживающийся процесс,
ведущий к тяжелому повреждению мембран.
К активным формам кислорода (АФК)
относятся супероксид (O2), синглетный
кислород, Н2О2 и радикал гидроксила
(ОН'). В организме человека и животных
первичным АФК служит супероксид,
возникающий при одноэлектронном
восстановлении молекулярного кислорода.
2.Окислительное фосфорилирование – это многоэтапный процесс, происходящий во внутренней мембране митохондрий и заключающийся в окислении восстановленных эквивалентов (НАДН и ФАДН2) ферментами дыхательной цепи и сопровождающийся синтезом АТФ.


Коэффициент Р/О – это отношение количества неорганического фосфата,
включенного в молекулу АТФ АТФ-синтазой, к количеству атомов кислорода,
включенного в молекулу Н2О, при переносе одной пары электронов по
дыхательной цепи.
3. Главная роль убихинона в организме – перенос электронов в мембране митохондрий. Благодаря этому он играет центральную роль в процессе окислительного фосфорилирования в клетке и образовании АТФ.
Гиповитаминоз
Благодаря тому, что убихинон способен синтезироваться в организме, выраженных симптомов его недостаточности не зафиксировано. Однако при некоторых патологиях, вследствие которых нарушается синтез убихинона, развивается анемия. Также возможно возникновение сердечной недостаточности вследствие нарушения работы миокарда и дистрофические изменения в скелетной мускулатуре.
Гипервитаминоз
Гипервитаминоз возникает только при повышенном введении убихинона. При этом возникают боли в области живота, нарушения стула и тошнота. Коэнзим Q является эффективным средством при лечении мышечной дистрофии (в том числе генетической ее формы) и сердечной недостаточности.

Билет 11
1 . Оксидазы
Тип реакций: перенос электронов и протонов водорода (дегидрирование) с одного субстрата на другой Акцептор водорода: кислород Продукт реакции: вода Кофакторы: медь, железо Название: субстрат + оксидаза ПРИМЕР: цитохром-с оксидаза Фермент процесса тканевого дыхания в митохондриях, где электроны, высвобождаемые из молекул различных субстратов при их полном окислении в клетке, переносятся на кислород с образованием метаболический воды О2 + 4Н+ + 4 е- → 2Н2О
Аэробные дегидрогеназы
Тип реакций: дегидрирование Акцептор водорода: кислород Продукт реакции: Н2О2 Кофермент (простетическая группа): FMN, FAD (производные вит. В2 - рибофлавина) Кофакторы: ионы металлов Кофакторы и коферменты – посредники в переносе электронов и протонов. В структуре кофермента именно витамин принимает и передает электроны, являясь, таким образом, «рабочей частью» кофермента. Название: субстрат + оксидаза ПРИМЕР: ксантиноксидаза (FAD, Mo2+, Fe3+)
ксантин + O2 + H2O → мочевая кислота + Н2О2 (реакция катаболизма пуриновых нуклеотидов)
Анаэробные дегидрогеназы
Тип реакций: дегидрирование Акцептор водорода – коферменты: -обычно NAD+ , NADP+ (производные вит. РР, или В3 – никотиновой кислоты) -реже FMN, FAD (производные вит. В2 – рибофлавина) Образуются восстановленные формы коферментов – NADH+H+, NADPH+H+, FADH2, FMNH2 «Рабочей частью» коферментов, принимающей электроны и протоны, являются витамины Название: субстрат + дегидрогеназа или редуктаза Название «субстрат+дегидрогеназа» подчеркивает важную роль процесса окисления субстрата Название «субстрат+редуктаза» подчеркивает важную роль процесса восстановления субстрата

2. Главные продукты реакции - это NADH+H+ и ацетил-СоА. NADH+H+ далее окисляется в дыхательной цепи, где энергия используется на синтез 3 моль АТР, а ацетил-СоА окисляется в цитратном цикле
3.
Дыхательный контроль – это прямое влияние электрохимического градиента на скорость движения электронов по дыхательной цепи (т.е. на величину дыхания). Величина градиента зависит от соотношения АТФ / АДФ, количественная сумма которых в клетке практически постоянна.
Разобщение дыхания и фосфорилирования Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из межмембранного пространства через мембрану в матрикс, минуя протонные каналы АТФ-синтазы. В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это явление называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается. В этом случае скорость окисления NADH и FADH2возрастает, возрастает и количество поглощённого кислорода, но энергия выделяется в виде теплоты, и коэффициент Р/О резко снижается.
Нарушения какого-либо этапа метаболизма, приводящие к прекращению синтеза АТФ, гибельны для клетки. Состояния, при которых синтез АТФ снижен, объединяют термином "гипоэнергетические". Причинами гипоэнергетических состояний могут быть голодание, гиповитаминозы В1, РР, В2; гипоксия. Гипоксия может возникнуть: при недостатке кислорода во вдыхаемом воздухе; при заболеваниях лёгких и нарушении лёгочной вентиляции. Причинами гипоксии могут быть также наследственные или приобретенные нарушения структуры гемоглобина. Частой причиной гипоэнергетических состояний могут быть нарушения процессов использования кислорода в клетках.
Причинами этих нарушений могут быть: - действие ингибиторов и разобщителей в ЦПЭ; - железодефицитные анемии; - снижение уровня гемоглобина и других железосодержащих белков (цитохромов, FeS-белков), в результате чего нарушаются перенос электронов и синтез АТФ; - наследственные дефекты ферментов ЦПЭ и цитратного цикла.
Билет 12
1.
2. ЦТК- центральная часть общего пути катаболизма, циклический биохимический процесс, в ходе которого ацетильные[en] остатки (СН3СО-) окисляются до диоксида углерода (CO2). При этом за один цикл образуется 2 молекулы CO2, 3 НАДН, 1 ФАДH2 и 1 ГТФ (или АТФ)[3]. Электроны, находящиеся на НАДН и ФАДH2, в дальнейшем переносятся на дыхательную цепь[2], где в ходе реакций окислительного фосфорилирования образуется АТФ.
Цикл протекает в матриксе митохондрий и представляет собой окисление молекулы ацетил-SКоА в восьми последовательных реакциях.
Цикл трикарбоновых кислот — это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей в организме, промежуточный этап между гликолизом и электронтранспортной цепью.
Функции ЦТК : 1. Энергетическая
генерация атомов водорода для работы дыхательной цепи, а именно трех молекул НАДН и одной молекулы ФАДН2,
синтез одной молекулы ГТФ (эквивалентна АТФ).
3.
Субстратное фосфорилирование – синтез АТФ из АДФ и фосфорной кислоты с использованием энергии высокоэнергетического субстрата. Этот способ синтеза АТФ не требует присутствия кислорода, т.е. происходит в анаэробных условиях.
Реакции субстратного фосфорилирования происходят:
1) в процессе гликолиза (макроэргические субстраты - 3-фосфо-глицерат и фосфоенолпируват);
2) с использованием креатинфосфата;
3) в одной реакции ЦТК, с использованием макроэргического субстрата – сукцинил-КоА.
Субстратное фосфорилирование служит для быстрой регенерации АТФ независимо от доступности акцепторов электронов для дыхательной цепи переноса, то есть в отсутствии кислорода. У человека в эритроцитах полностью отсутствует аэробное дыхание и вся энергия генерируется исключительно за счёт субстратного фосфорилирования гликолиза. Также субстратное фосфорилирование способствует более полному использованию энергии окисляемых веществ.



Билет 13
В нашем организме окисление может происходить двумя способами: 1. Путем отнятия водорода от окисляемого субстрата: сюда относятся МИТОХОНДРИАЛЬНОЕ ОКИСЛЕНИЕ и ВНЕМИТОХОНДРИАЛЬНОЕ ОКИСЛЕНИЕ ОКСИДАЗНОГО ТИПА. 2. Путем присоединения кислорода к окисляемому субстрату - так происходит внемитохондриальное ОКИСЛЕНИЕ ОКСИГЕНАЗНОГО
ТИПА (старое название - МИКРОСОМАЛЬНОЕ окисление).
Механизм сопряжения окисления и фосфорилирования
Строение АТФ-синтеза

Синтез АТФ
Синтез АТФ происходит за счет энергии, высвобождающейся при прохождении протонов через каналы комплекса Fo. Протоны из межмембранного пространства достигают своих центров связывания на с-субъединицах (Fo ) через входной полуканал и прикрепляются к аспартату, нейтрализуя его отрицательный заряд. Лишенный заряда белок способен вращаться вокруг своей оси, доставляя протоны к другому полуканалу, который направлен уже в матрикс митохондрий. Поскольку концентрация ионов H+ в матриксе низка, то они легко отрываются от аспартата и уходят внутрь, а оставшийся заряженный аспартат не позволяет ротору вращаться в обратном направлении.
При связывании ионов H+ происходит поворот комплекса c-субъединиц и связанной с ним γ-субъединицы другого компонент. Стержень γ–субъединицы проворачивается и при повороте на 120° она поочередно вступает в контакт с каталитическими β-субъединицами, что меняет их функционирование. β-субъединицы могут находиться в трех конформациях, выполняющих разную функцию: • loose, L – удерживает АДФ и ион фосфата, • tight, T– "прижимает" молекулы АДФ и фосфат-иона, т.е. происходит синтез АТФ, • open, O – в этом состоянии субъединица высвобождает АТФ и захватывает АДФ и ион фосфата.
Цтк для укороченной цепи
Известны два субстрата. Главным из них является сукцинат (янтарная кислота). Сукцинатдегидрогеназа – это комплекс II, который в укороченном варианте цепи является начальным звеном окисления. В составе комплекса – простетическая группа ФАД и FeSII. От ФАД.Н2 два атома водорода переносятся на KoQ. Значит, первое звено, которое имеется в полной цепи - исключается. Перепад окислительно-восстановительного потенциала между ФАД и KoQ невелик. Поэтому переноса H+ в межмембранное пространство в этой точке не происходит. DmH+ также создается, но меньший, чем в полной цепи. Значит, меньше и эффективность фосфорилирования - коэффициент Р/О=2. Аналогичным образом окисляется и второй субстрат – ацил-КоА (активная форма любой жирной кислоты)
Ферменты– это биокатализаторы преимущественно белковой природы (иРНК тоже имеют ферментативную активность), которые принимают участие в химических реакциях в организме. Значение ферментов: ферменты принимают участие в большинстве процессов проис ходящих в организме реакциях синтеза и распада веществ, процессах переваривания и всасывания, освобождения энергии, обеспечивают координацию биохимических реакций.
Коферменты – это небелковые компоненты сложных ферментов, которые проявляют высокую химическую активность и входят в состав активных центров сложных ферментов. Классификация коферментов. 1. По химической природе: 1) витаминные; 2) витаминоподобные; 3) невитаминные. 2. По механизму действия: 1) переносчики атомов водорода, электронов и протонов. 2) переносчики отдельных химических групп. Энергетический эффект какого-либо биологического вещества выражают количеством АТФ, которое можно получить в данном процессе. Расчет энергетического эффекта биохимических процессов, протекающих в анаэробных и аэробных условиях, следует производить по-разному.
Билет 14
Катаболизм (диссимиляция ) - это энергетический обмен, часть метаболизма, распада сложных веществ на более простые (или окисления вещества), который протекает с освобождением энергии в виде тепла и в виде молекулы АТФ, универсального источника энергии всех биохимических процессов. Схема катаболизма пищевых веществ, его этапы, пути и процессы На схеме наглядно представлен катаболизм пищевых веществ, который состоит из 3-х основных этапов (стадий), первый и второй этапы относятся к специфическим путям катаболизма, а третий этап относится к общему пути катаболизма.
ATP (АТФ) - это аденозинтрифосфорная кислота (нуклеотид), универсальный источник и переносчик энергии, который участвует во всех биохимических процессах.
NAD (НАД) - никотинамидадениндинуклеотидфосфата, является коферментом, переносит электроны из одной реакции в другую в метаболизме.
NADH (НАД*Н) - востановленная форма NAD.
Пируват - это соли пировиноградной кислоты, конечный продукт в процессе гликолиза

Этапы катаболизма |
Протекание процессов катаболизма |
Энергетическая ценность |
1-й этап катаболизма (подготовительный) расщепление больших макромолекул на простые субьединицы |
На первом этапе происходит расщепление пищевых веществ (белки, полисохариды, жиры) до мономеров. У многоклеточных организмов это осуществляется в пищеварительном тракте (у простейших в лизосомах при самообновленнии клеток) под воздействием соответствующих ферментов, после чего полученные мономеры всасываются в кровь (моносахариды аминокислоты) и в лимфу (жирные кислоты). |
Небольшое количество энергии рассеивается ввиде тепла |
2-й этап катаболизма (бескислородный) расщепление простых субьединиц на ацетил-СоА, сопровождающийся образованием ограниченного количества АТP и NADH |
На втором этапе все пищевые продукты которые поступают в клетку из крови образуют ацетилкоэнзим А (ацетил-СоА). Это соединение, а также другие ферменты, включающие в себя КоА, являются ключевыми звеньями множества разнообразных биохимических реакций. — При гликолизе моносахариды образуют пировиноградную кислоту. — При катаболизме аминокислот образуются ацетил-CoA, пируват, другие кетокислоты — При β-окислении жирных кислот образуется ацетил-CoA Второй этап происходит в цитозоли и митохондрии. |
При расщеплении глюкозы 60% выделившейся энергии дает тепло, 40% идет на синтез 2х молекул ATP, эта часть энергии запасается. |
3-й этап катаболизма (кислородный) при полном окислении ацетил-СоА до H2O и CO2 образуется большое количество NADH, что обеспечивает синтез большого количества ATP при переносе электронов |
Третий этап и его реакции проходят в митохондриях. Ацетил-CoA участвует в реакциях цикла лимонной кислоты (цикл трикарбоновых кислот Кребса), там углероды окисляются до углекислого газ CO2. Происходит полное окисление ацетильной группы ацетил-СоА до Н2O и СO2, при этом большое количество электронов и протонов запасается на молекулах NADH (процесс " |
При окислении 2х молекул кислоты образуется 36 молекул ATP |
2.
Энергия, накопленная в одних макроэргах, может передаваться от одной молекулы к другой. Для некоторых макроэргов характерным является то, что накопленная в них энергия может непосредственно использоваться для совершения различных видов работы. Например, 1,3-дифосфоглицерат, фосфоенолпируват и креатинфосфат принимают участие в реакциях субстратного фосфорилирования, в которых эти субстраты передают свою макроэргическую связь на АДФ с образованием АТФ. Креатинфосфат находится в больших концентрациях в мышцах, где выполняет функцию депо энергии для мышечного сокращения. Карбамоилфосфат используется в клетках печени для синтеза мочевины. АТФ является универсальной энергетической валютой клетки, потому что только это соединение способно непосредственно отдавать свою энергию, накопленную в макроэргических связях, на совершение любого вида работы или энергозависимого процесса.
3. В большинстве эукариотических клеток синтез АТФ происходит внутри митохондрии, а основные потребители АТФ расположены вне её. С другой стороны, в матриксе митохондрий должна поддерживаться достаточная концентрация АДФ. Эти заряженные молекулы не могут самостоятельно пройти через липидный слой мембран. Внутренняя мембрана непроницаема для заряженных и гидрофильных веществ, но в ней содержится определённое количество транспортёров, избирательно переносящих подобные молекулы из цитозоля в матрикс и из матрикса в цитозоль.
В мембране есть белок АТФ/АДФ-антипортер, осуществляющий перенос этих метаболитов через мембрану.

Потоки различных веществ (АТФ, АДФ, Н3РО4, Са2+) проходят через специфические транспортёры, при этом затрачивается энергия электрохимического потенциала мембраны.
Молекула АДФ поступает в митохондриальный матрикс только при условии выхода молекулы АТФ из матрикса.
Движущая сила такого обмена - мембранный потенциал переноса электронов по ЦПЭ. Расчёты показывают, что на транспорт АТФ и АДФ расходуется около четверти свободной энергии протонного потенциала. Другие транспортёры тоже могут использовать энергию электрохимического градиента. Так переносится внутрь митохондрии неорганический фосфат, необходимый для синтеза АТФ. Непосредственным источником свободной энергии для транспорта Са2+ в матрикс также служит протонный потенциал, а не энергия АТФ.
Билет №15