

Вопрос 89
Циклические АМФ и ГМФ как вторичные посредники. Активация протеинкиназ и фосфорилирование белков, ответственных за проявление гормонального эффекта.
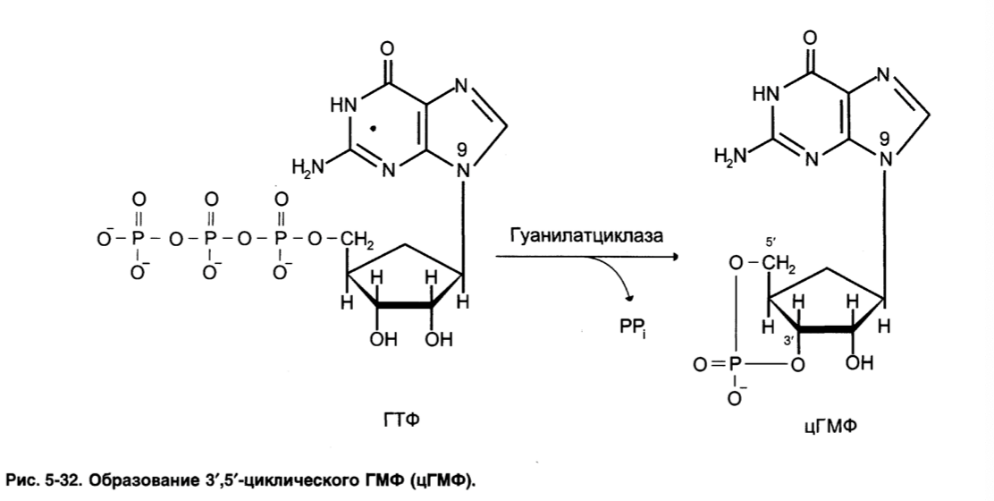
Гуанилатциклаза катализирует образование цГМФ из ГТФ, одного из важных
посредников внутриклеточной передачи сигнала. Она находится в клетке, в мембранносвязанном и в цитозольном состоянии.
Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ – ключевой фермент аденилатциклазной системы передачи сигнала. Аденилатциклаза обнаружена во всех типах клеток.

Активация протеинкиназы А (ПКА)
Молекулы цАМФ могут обратимо соединяться с регуляторными субъединицами ПКА.
Активная протеинкиназа А фосфорилирует специфические белки по серину и треонину, в результате изменяются конформация и активность фосфорилированных белков, это приводит к изменению скорости и направления регулируемых ими процессов в клетке.
Концентрация цАМФ зависит от соотношения активностей ферментов аденилатциклазы и фосфодиэстеразы.
Регуляция синтеза и распада гликогена в печени глюкагоном и адреналином.
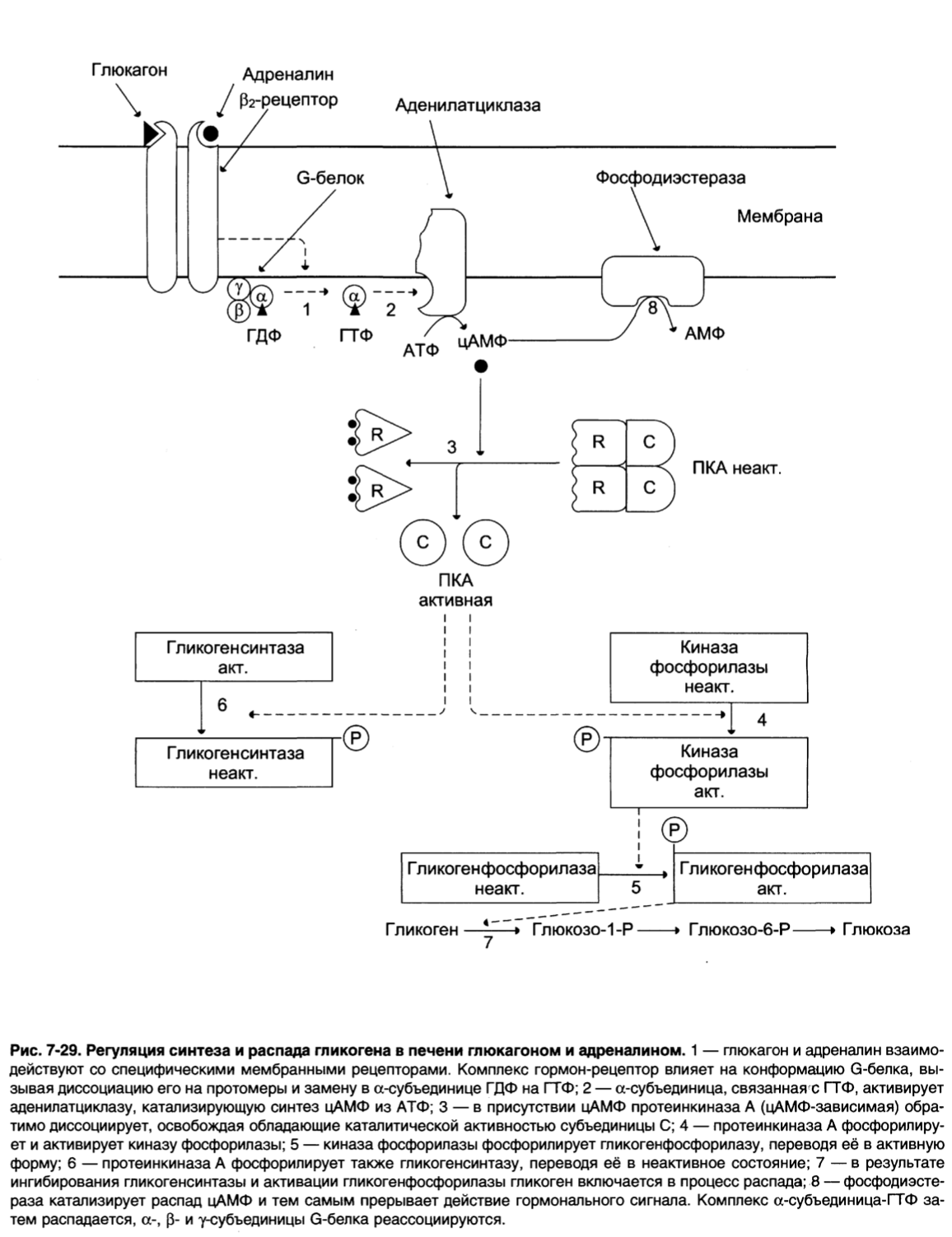
1 – глюкагон и адреналин взаимодействуют со специфическими мембранными рецепторами. Комплекс гормон-рецептор влияет на конформацию G-белка, вызывая диссоциацию его на протомеры и замену в α-субъединице ГДФ на ГТФ;
2 – α-субъединица, связанная с ГТФ, активирует аденилатциклазу, катализирующую синтез цАМФ из АТФ;
3 – в присутствии цАМФ протеинкиназа А (цАМФ-зависимая) обратимо диссоциирует, освобождая обладающие каталитической активностью субъединицы С;
4 – протеинкиназа А фосфорилирует и активирует киназу фосфорилазы;
5 – киназа фосфорилазы фосфорилирует гликогенфосфорилазу, переводя её в активную форму;
6 – протеинкиназа А фосфорилирует также гликогенсинтазу, переводя её в неактивное состояние;
7 – в результате ингибирования гликогенсинтазы и активации гликогенфосфорилазы гликоген включается в процесс распада;
8 – фосфодиэстераза катализирует распад цАМФ и тем самым прерывает действие гормонального сигнала. Комплекс α-субъединица-ГТФ затем распадается, α-, β- и у-субъединицы G-белка реассоциируются.
Активация протеинкиназы с
Повышение концентрации Са2+ в цитозоле клетки увеличивает скорость взаимодействия Са2+ с неактивным цитозольным ферментом протеинкиназой С (ПКС) и белком кальмодулином, таким образом сигнал, принятый рецептором клетки, раздваивается.
Связывание протеинкиназы С с ионами кальция позволяет ферменту вступать в кальций-опосредованное взаимодействие с молекулами "кислого" фосфолипида мембраны, фосфатидилсерина (ФС). Диацилглицерол, занимая специфические центры в протеинкиназе С, ещё более увеличивает её сродство к ионам кальция.
Регуляция синтеза и распада гликогена в печени адреналином и Са2+.
ФИФ2 – фосфатидилинозитолбисфосфат;
ИФ3 – инозитол-1,4,5-трифосфат;
ДАГ – диацилглицерол;
ЭР – эндоплазматический ретикулум;
ФС – фосфодитилсерин.
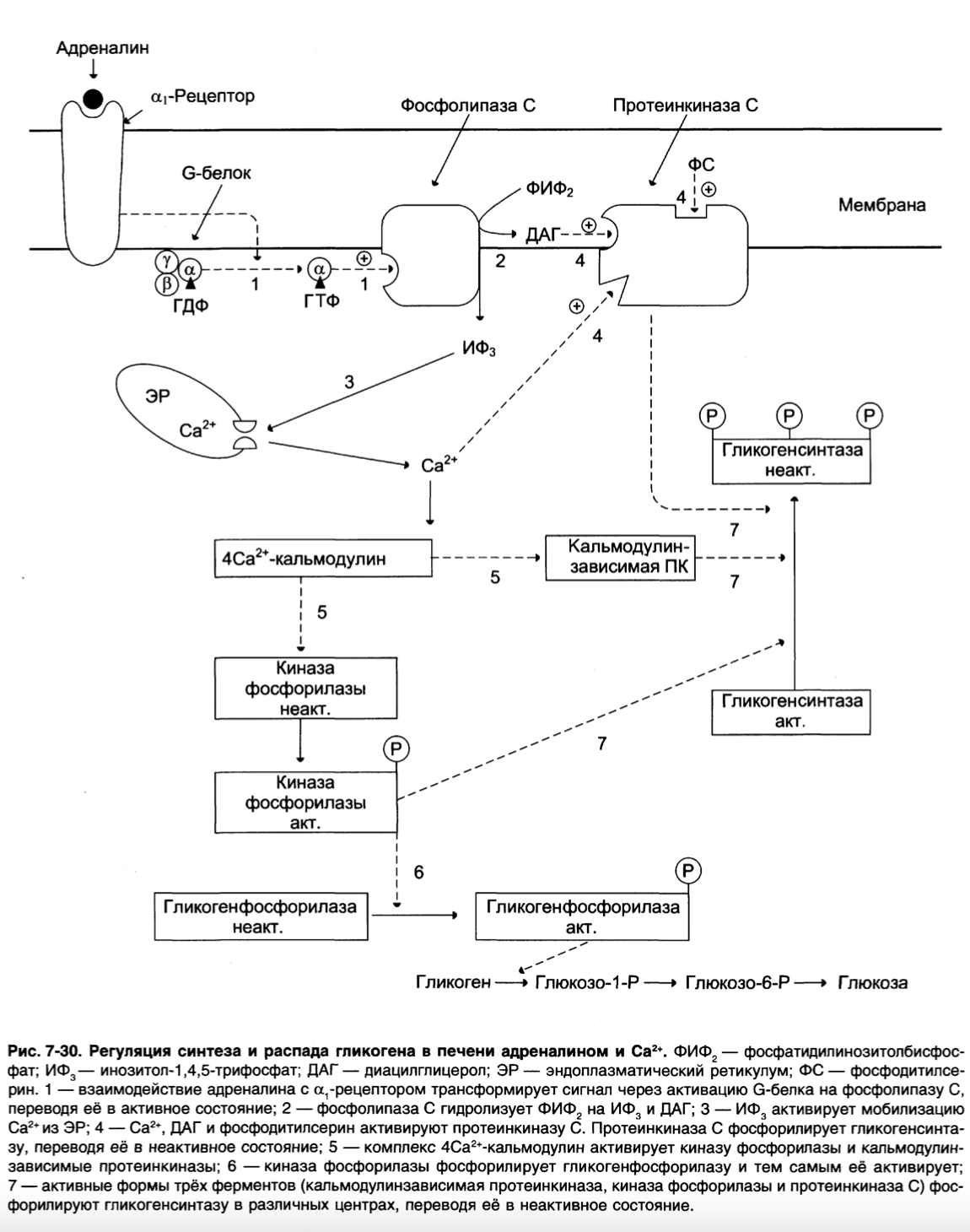
1 – взаимодействие адреналина с α1-рецептором трансформирует сигнал через активацию G-белка на фосфолипазу С, переводя её в активное состояние;
2 – фосфолипаза С гидролизует ФИФ2 на ИФ3, и ДАГ;
3 – ИФ3, активирует мобилизацию Са2+ из ЭР;
4 – Са2+, ДАГ и фосфодитилсерин активируют протеинкиназу С.
Протеинкиназа С фосфорилирует гликогенсинтазу, переводя её в неактивное состояние;
5 – комплекс 4Са2+-кальмодулин активирует киназу фосфорилазы и кальмодулинзависимые протеинкиназы;
6 – киназа фосфорилазы фосфорилирует гликогенфосфорилазу и тем самым её активирует;
7 – активные формы трёх ферментов (кальмодулинзависимая протеинкиназа, киназа фосфорилазы и протеинкиназа С) фосфорилируют гликогенсинтазу в различных центрах, переводя её в неактивное состояние.