

Fiza_2
.pdfЗанятие № 2
1.
Нейро́н, или невро́н (от др.-греч. νεῦρον — волокно, нерв) — структурно-функциональная единица нервной системы. Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов
Классификация:
1.В зависимости от нейромедиатора, используемого для передачи:
-холинергические – медиатор ацетилхолин (АХ);
-адренергические – норадреналин (НА);
-серотонинергические – серотонин (СТ);
-глицинергические – аминокислота глицин (ГЛИ);
-ГАМК-ергические – гамма-аминомасляная кислота (ГАМК);
-дофаминергические – дофамин (ДА);
-пептидергические – медиаторами являются нейропептиды ( вещество Р, опиоидный пептид в-эндорфин и др.)
2. По форме (до 80 вариантов нейронов):
- звёздчатые - пирамидальные,
- грушевидные - веретеновидные - паукообразные - овальные
3. Функциональная (в зависимости от выполняемой функции и места в рефлекторной
дуге): |
|
- рецепторные (чувствительные, афферентные нейроны с помощью дендритов вос |
- |
принимают воздействия внешней или внутренней среды, ге нерируют нервный импульс и передают его другим типам нейронов , встречаются только в спинальных ганглиях и
чувствительных ядрах черепномозговых нервов) |
|
||
- |
эффекторные |
(эфферентные нейроны передают возбуждение на ра |
бочие органы, |
например, мышцы или железы, располагаются в передних рогах спинного мозга и |
|||
вегетативных нервных ганглиях) |
|
||
- |
вставочные |
(ассоциативные нейроны располагаются между рецепторными и |
|
эффекторными нейронами; по количеству их больше всего, особенно в ЦНС)
- секреторные (нейросекреторные клетки, специализированные нейроны, по своей функции напоминающие эндокринные клетки , синтезируют и выделяют в кровь нейрогор моны, расположены в гипоталамической области головного мозга, регулируют
4. По физиологической роли:
-возбуждающие
-тормозные
5. По количеству и характеру отростков:
-униполярные (имеющие один отросток)
-псевдоуниполярные (от тела отходит один отросток, который затем делится на 2 ветви)
-биполярные (2 отростка, один дендритоподобный, другой аксон)
-мультиполярные (имеют 1 аксон и много дендритов)
Физиологические функции:
1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;

2) проводимость – способность передавать нервные возбуждение в виде потенциала действия от места раздражения по всей длине; 3) рефрактерность (устойчивость) – свойство временно резко снижать возбуди мость в процессе возбуждения.
Нервная ткань имеет самый короткий рефрактерный период. Значение рефрактерности – предохранять ткань от перевозбуждения, осуществляет ответную реакцию на биологически значимый раздражитель; 4) лабильность – способность реагиро вать на раздражение с определенной скоростью.
Лабильность характеризуется максимальным числом импульсов возбуждения за определенный период времени (1 с) в точном соответствии с ритмом наносимых раздражений.
2.
На рис. 31 показаны три состояния нейрона.
Рис. 31. Три состояния нейрона. А. Нейрон в покое с ПП ( -65 мВ); Б. Нейрон в состоянии возбуждения с ВПСП (-45 мВ) и с переходом его в ПД; В. Нейрон в заторможенном состоянии с ТПСП (-70 мВ) (Гайтон, 2008)
При возбуждении нейронов потребление кисл орода возрастает. Источником энергии служит в основном глюкоза крови, собственных небольших запасов гликогена хватает лишь на 3-5 мин работы нейрона. Передача сигнала в химических синапсах ЦНС подобна таковой в нервно -мышечном синапсе, однако имеет ряд отл ичительных особенностей, основные из которых приводятся ниже.
Для возбуждения нейрона (возникновения ПД) необходимы потоки афферентных
импульсов и их взаимодействие. Одна везикула (квант медиатора) содержит 1 —10 тыс.
молекул медиатора. Один ПД, пришедший в пресинаптическое окончание, обеспечивает выделение 200—300 квантов медиатора; при этом возникает небольшой ВПСП — около 0,05 мВ (миниатюрный ВПСП).
Необходимо учесть, что одновременно могут возникать не только возбуждающие, но и тормозные потенциалы. На нейронах одновременно присутствуют возбуждающие и тормозные синапсы (рис. 32).
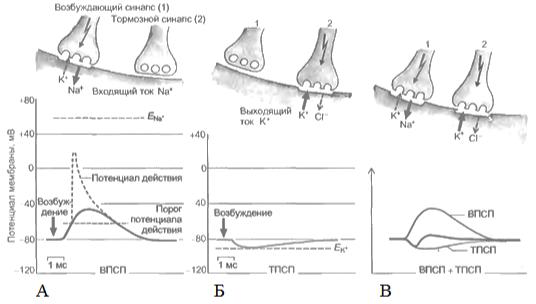
Рис. 32. А - Процессы освобождения медиатора, поступления ионов натрия и формирования
ВПСП в возбуждающем синапсе и потенциала действия ПД; Б |
- Процесс формирования |
тормозного постсинаптического потенциала (ТПСП) в тормозном синапсе после поступления медиатора и выхода ионов калия или входа ионов хлора; В – Процесс суммации ТПСП и ВПСП на постсинаптической мембране нейрона. В верхней части рисунка показаны процессы, развивающиеся в синапсах (возбуждающий, тормозной), а в нижней – графическое отображение этих процессов (ВПСП,ТПСП и ПД) (Камкин, 2004)
Пороговый потенциал нейрона равен 5 —10 мВ, поэтому ясно, что для возбуждения нейрона требуется некоторое множество импульсов. Выброс медиатора из нервного
окончания обеспечивает входящий в деполяризованную терминаль Са |
2+, причем |
количество медиатора прямо пропорционально входу ионов Са 2+, при э том четыре иона Са2+ обеспечивают выброс одного кванта медиатора. При поступлении импульсов к нейрону-мишени по входам в результате пространственной суммации ВПСП возникает ПД нейрона-мишени.
ВПСП возникает вследствие суммарного тока в клетку и из клетки ч ерез ионные каналы различных ионов согласно электрохимическому градиенту. Поступивший в пресинаптическое окончание Са2+ удаляется в среду с помощью Са-насоса.
Прекращение действия выделившегося в синаптическую щель медиатора осуществляется частично посредством обратного захвата его пресинаптическим окончанием, частично — с помощью разрушения специальными ферментами. Например, норадреналин расщепляется МАО и катехолметилтрансферазой, ацетилхолин гидролизуется
ацетилхолинтрансферазой, имеющейся в синаптическо |
й щели и встроенной в |
|
постсинаптическую мембрану. Прекращение действия избытков медиатора на |
||
постсинаптическую мембрану предотвращает десенситизацию |
— снижение |
|
чувствительности постсинаптической мембраны к действующему медиатору.
В возникновении ПД в нейро нах принимают участие ионы Сa 2+, ток которых в клетку более медленный, чем ток Na+, играющий главную роль в формировании ПД-нейронов. В частности, в дендритах клеток Пуркинье мозжечка выявлены не только быстрые натриевые потенциалы, но и медленные кальциев ые. В телах некоторых нервных клеток потенциал действия создается преимущественно за счет Са2+.
Место возникновения ВПСП, вызывающих ПД нейрона , — тело нейрона , поскольку постсинаптические мембраны этих синапсов располагаются в непосредственной близости от аксонного холмика, где начинается развитие ПД. Близость ВПСП, возникающих в теле
нейрона, к аксонному холмику обеспечивает их участие в механизмах генерации П |
Д. |
Однако подавляющее большинство синапсов (в частности, в коре большого мозга, |
|
согласно расчетам, 98%) находится на дендритах. Площадь мембраны тела нейронов занята синапсами на 40%, дендритов — на 75%. Синапсов в коре большого мозга в 40 тыс. раз больше, чем самих нейронов.
Место возникновения ПД нейрона. Формирование ПД начинается на мембране аксонного холмика (генераторный пункт нейрона). Синапсы на нем отсутствуют, возбудимость мембраны аксонного холмика в 3 —4 раза превосходит возбудимость мембраны тел а нейрона, что объясняется более высокой (примерно в 7 раз) концентрацией Na - каналов на аксонном холмике (на нем отсутствует миелиновая оболочка). ВПСП, возникающие в любых участках сомы нейрона суммируются, их суммарное электрическое поле достигает аксонного холмика и вызывает его деполяризацию до КП, в результате чего начинает развиваться ПД на аксонном холмике.Это связано с тем, что постоянная длины мембраны тела нейрона (расстояние, на котором исходная амплитуда ВПСП уменьшается на 37%) составляет 1-2 мм, а диаметр тела нейрона — всего лишь 10 -80 мкм. Поэтому электрическое поле начального возбуждения аксонного холмика обеспечивает деполяризацию тела нейрона до КП, в результате чего возникает ПД нейрона, который проводится по аксону к другой клетке.
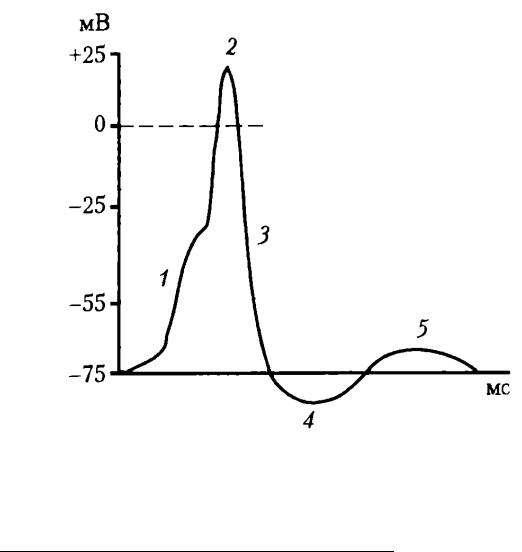
На восходящей части ПД нейрона регистрируется перегиб на уровне 30— 40 мВ (задержка в развитии ПД), вследствие того , что возбудимость мембраны тела нейрона в 3 —4 раза ниже возбудимости мембраны аксонного холмика, что также является подтверждением описанного механизма формирования ПД нейрона (рис. 33).
Рис. 33. Типичный ПД нейронов ЦНС: 1 – деполяризация; 2 – инверсия; 3 – реполяризация; 4 – следовая гиперполяризация; 5 – следовая деполяризация
Таким образом, ПД нейрона |
— это единый одновременный процесс воз буждения |
аксонного холмика и тела нейрона.
Роль дендритов в возникновении возбуждения нейрона. Дендритные синапсы называют модуляторами. Это связано с тем, что они удалены на значительное расстояние от генераторного пункта нейрона — аксонного холмика. Поэтому суммарное электрическое
поле их ВПСП не может вызвать достаточную деполяризацию аксонного холмика и обеспечить возникновение ПД, а определяет лишь возбудимость нейрона.
3.
Миелиновые волокна:
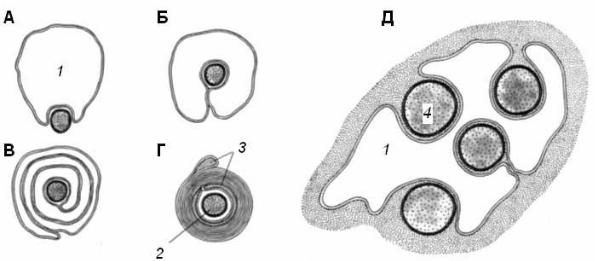
Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
|
|
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А |
– Г); |
соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000) |
|
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно |
|
|
|
Безмиелиновые волокна:
Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития.
Влеммоцит по гружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
Механизм проведения возбуждения по безмиелиновым нервным волокнам:
Всостоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит
реверсия заря да (рис. 2, А). На границе возбужденного и невозбужденного участка
начинает протекать электрический ток (рис. 2, |
Б). Электрический ток раздражает |
ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
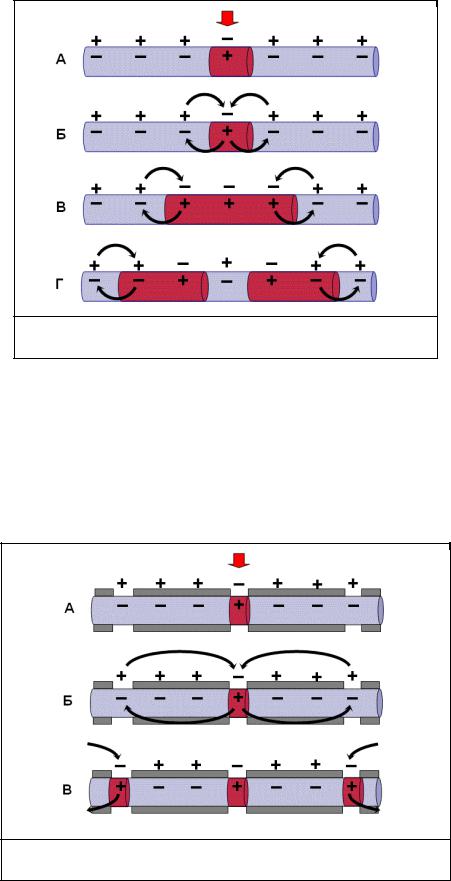
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
Механизм проведения возбуждения по миелиновым нервным волокнам:
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье.
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны ( рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
4.
Нервные волокна (neurofibrae) – это отростки нервных клеток, окруженные оболочками из нейроглиальных клеток.
Существуют различные классификации нервных волокон.

По наличию (или отсутствию) миелиновой оболочки нервные волокна делят на миелиновые
и безмиелиновые. |
|
Классификация нервных волокон по Эрлангеру |
– Гассеру основана на исследовании |
составного потенциала действия смешанного пери |
ферического нерва (например, |
седалищного). После электрической стимуляции такого нерва регистрируется целый спектр потенциалов действия, отражающий наличие в нерве волокон с разными скоростями проведения импульса. Согласно этой классификации выделяют три основных типа волокон
– А, В, С, соответствующие трем волнам составного потенциала действия. Волокна типа А подразделяются на 4 подтипа – α, β, γ, δ. Данная классификация учитывает как чувствительные, так и двигательные волокна.
Классификация нервных волокон по Ллойду– Ханту учитывает диаметр волокна и скорость проведения импульса, но применяется только для чувствительных волокон. Согласно этой классификации выделяют четыре класса волокон– I,II,III,IV. ВолокнаIкласса подразделяют на 2 группы – а иb. Эти кл ассификации не противоречат друг другу и представлены в сводной таблице.
Тип |
Тип волокна |
Диаметр |
Скорость |
Функция, тип обслуживаемого |
|
волокна |
(только |
волокна |
(м/с) |
рецептора |
|
|
чувствительные) |
(мкм) |
|
|
|
|
|
|
|
|
|
Чувствительные нервные волокна |
|
|
|
||
|
|
|
|
|
|
Аα |
Ia, Ib |
13 – 20 |
80 – 120 |
Первичные афферентные волокна |
|
|
|
|
|
мышечных веретен и от |
|
|
|
|
|
сухожильных органов Гольджи. |
|
|
|
|
|
|
|
Аβ |
II |
6 – 12 |
35 – 75 |
Вторичные афферентнфе волокна |
|
|
|
|
|
мышечных веретен, кожные |
|
|
|
|
|
механорецепторы |
|
|
|
|
|
|
|
Аδ |
III |
1 – 5 |
5 – 30 |
Кожные механорецепторы, |
|
|
|
|
|
терморецепторы, ноцицепторы |
|
|
|
|
|
|
|
С |
IV |
0,2 – 1,5 |
0,5 – 2 |
Кожные механорецепторы, |
|
|
|
|
|
терморецепторы, ноцицепторы |
|
|
|
|
|
(немиелинизированные волокна) |
|
|
|
|
|
|
|
Двигательные нервные волокна |
|
|
|
||
|
|
|
|
|
|
Аα |
− |
12 – 20 |
72 – 120 |
Экстрафузальные волокна |
|
|
|
|
|
скелетных мышц |
|
|
|
|
|
|
|
Аγ |
− |
2 – 8 |
12 – 48 |
Интрафузальные |
мышечные |
|
|
|
|
волокна |
|
|
|
|
|
|
|
В |
− |
1 – 3 |
6 – 18 |
Преганглионарные вегетативные |
|
|
|
|
|
волокна |
|
|
|
|
|
|
|
С |
− |
0,2 – 2 |
0,5 – 2 |
Постганглионарные вегетативгые |
|
|
|
|
|
(немиелинизированные волокна) |
|
|
|
|
|
|
|
5.
Закон анатомо -физиологической целостности: проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных

наркотических средств, сдавливания, а также порезами и повре ждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
6.
Явление парабиоза открыто русским физиологом Н.Е.Введенским в 1901 г. при изучении возбудимости нервно -мышечного препарата. Состояние парабиоза могут вызвать различные воздействия – сверхчастые, сверхсильные стимулы, яды, лекарства и другие воздействия как в норме, так и при патологии. Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента), то лабильность т акого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности и было названо Н. Е. Введенским парабиозом. Состояние парабиоза возбудимой ткани возникает под влиянием сильных раздражителей и характеризуется фазными нарушениями проводимости и возбудимости. Работы Н.Е.Введенского по парабиозу сыграли важную роль в развитии нейрофизиологии и клинической медицины, показав единство процессов возбуждения, торможения и покоя, изменили господствовавший в физиологии закон силовых отношений, согласно которому реакция тем больше, чем сильнее действующий раздражитель.
Выделяют 3 фазы
1) Примум (уравнительная - по И.П. Павлову; провизорная, трансформирующая - по Н.Е.Введенском): характеризуется снижением возбудимости и повышением лабильности. происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители
ответная реакция меньше, а на частые раздражители |
—больше. При действии |
парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотически й участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызва нной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т.е. 25 импульсов из 100), то амплитуда ответной реакции
становится такой же, как на редкие раздражители (25Гц) |
—происходит ура внивание |
ответной реакции. |
|
2) Оптимум (парадоксальная фаза) : возбудимость достигает максимума, лабильность начинает снижаться. В третью фазу (пессимум) возбудимость и лабильность снижаются параллельно и развивается 3 стадии парабиоза. Характеризуется извра щенным реагированием – сильные раздражения вызывают меньший ответ, чем умеренные. Происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно
меньше, т. к. часты |
е раздражители еще больше снижают лабильность, удлиняя фазу |
|
абсолютной рефрактерности. Следовательно, наблюдается парадокс |
— на редкие |
|
раздражители ответная реакция больше, чем на частые. |
|
|
3) Тормозная фаза: |
лабильность снижается до такой степени, что и редкие, и частые |
|
раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние. Ни сильные, ни умеренные раздражения не вызывают видимой реакции, в ткани развивается торможение. Парабиоз — явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности.

Однако, при действии сильных раздражителей за тормозной стадией может наступить полная потеря возбудимости и проводимости, а в дальнейшем – гибель ткани. Медицинское значение: явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ вязано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
7.
Закон двустороннего проведения возбуждения: нервное волокно проводит нервные
импульсы в двух направлениях – центростремительно и центробежно. В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
8.
Закон изолированного проведения возбуждения: существует ряд особенностей
распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах. В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе. В мякотных нервных волокнах роль изолятора выполняет миелиновая
оболочка. За счет миелина увеличивается удельное сопротивление и происходит
уменьшение электрической емкости оболочки. В безмякотных нервных волокнах
возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значител ьно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
9.
Физиологические свойства поперечнополосатых мышц:
-возбудимость (ниже, чем в нервном волокне, что объясняется низкой величиной мембранного потенциала)
-низкая проводимость, порядка 10–13 м/с
-рефрактерность (занимает по времени больший отрезок, чем у нервного волокна)
-лабильность
-сократимость (способность укорачиваться или развивать напряжение)
Физиологические функции поперечнополосатых мышц:
-двигательная (динамическая и статическая)
-обеспечения дыхания
-мимическая
-рецепторная
-депонирующая
-терморегуляторная
10.
При световой микроскопии было замечено, что в момент сокращения ширина А -диска не уменьшается, а Iдиски и Н -зоны саркомеров суживаются. При электронной, микроскопии было установлено, что длина нитей актина и миозина в момент соскращения не изменяется. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей. Согласно этой теории , мышца укорачивается в результате движения тонких актиновых нитей в промежутки между
миозиновыми. Это приводит к укорочению каждого саркомера |
, образующего |
миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное
состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя (гребковые движения). Но это последний этап всего сократительного механизма.
Сокращение начинается с того, что в области концевой пластинки двигательного нерва возникает ПД. Он с большой скоростью распространяется по сарколемме и переходит с неѐ по, системе поперечных трубочек СР, на продольные трубоч ки и цистерны. Возникает деполяризация мембраны цистерн и из них в саркоплазму высвобождаются ионы кальция. На нитях актина расположены молекулы еще двух белков – тропонина и тропомиозина. При низкой (менее 10-8 моль) концентрации кальция, т.е. в состоянии покоя, тропомиозин блокирует присоединение мостиков миозина к нитям актина. Когда ионы кальция начинают выходить из СР, молекула тропонина изменяет свою форму таким образом, что освобождает активные центры актина от тропомиозина. К этим центрам присоединя ются головки миозина и начинается скольжение за счет ритмического прикрепления и разъединения поперечных мостиков с нитями актина. При этом головки ритмически продвигаются по нитям актина к Z -мембранам. Для полного сокращения мышцы необходимо 50 таких циклов.
Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением . Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са -насос (фермент Са - АТФаза). Ионы кальция вновь закачиваются в цистерны саркоплазматического ретикулума и их концентрация падает ниже 10 -8 моль. Молекулы тропонина приобретают исходную форму и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от них , и мыш ца за счет эластичности приходит в исходное расслабленное состояние.
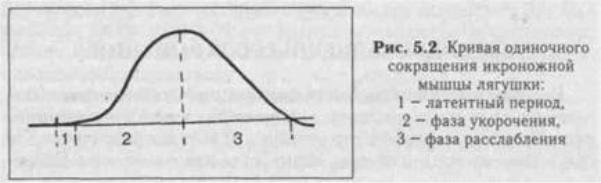
11.
При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода: 1) Латентный период : это время от момента нанесения раздражения до начала
сокращения. Его длительность около 1 -2 мс. Во время латентного периода генерируется и распространяется ПД, происходит высвобождение кальция из СР, взаимодействие актина с миозином и т.д.
2) Период укорочени я: в зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек., 3) Период расслаблени: его длительность нес колько больше, чем укорочения (0,05 -0.06 сек.)