

Общая микробиология
.pdf201
родов, около 2284 видов и свыше 3000 неклассифицированных еще вирусов.
В 1971 г. американский биохимик, молекулярный биолог и вирусолог лауреат Нобелевской премии Д. Балтимор предложил классификацию вирусов, основанную на типе геномной нуклеиновой кислоты и способе её репликации. Классификация вирусов по Балтимору включает в себя следующие группы:
I группа – вирусы, содержащие двухцепочечную (дц) ДНК и не имеющие РНК-стадии (например, герпесвирусы, поксвирусы).
II группа – вирусы, содержащие одноцепочечную (оц) ДНК (например, парвовирусы).
III группа – вирусы, содержащие двухцепочечную (дц) РНК (например, ротавирусы).
IV группа – вирусы, содержащие одноцепочечную (оц) плюс-РНК (например, пикорнавирусы, флавивирусы).
V группа – вирусы, содержащие одноцепочечную (оц) РНК негативной или двойной полярности (например, ортомиксовирусы, филовирусы).
VI группа – вирусы, содержащие одноцепочечную (оц) плюс-РНК и имеющие в процессе жизненного цикла стадию синтеза ДНК на матрице РНК (например, ретровирусы).
VII группа – вирусы, содержащие двухцепочечную (дц) ДНК и имеющие в процессе жизненного цикла стадию синтеза ДНК на матрице РНК (например, вирус гепатита В).
Современная классификация вирусов объединяет классификацию ICTV и классификацию по Балтимору. При этом учитываются такие свойства вирусов как морфология вирионов (размер и форма, тип симметрии, количество капсомеров), физико-химические свойства (молекулярная масса, плавучая плотность, устойчивость к физическим и химическим факторам), структура генома (тип нуклеиновой кислоты, размер, наличие сегментов, количество нитей, форма молекулы нуклеиновой кислоты, полярность), количество и свойства структурных и неструктурных белков, наличие липидов, генетическое сходство с другими вирусами, наличие суперкапсидной оболочки, таксономическая принадлежность организма – хозяина.
6.8. Жизненный цикл вирусов
Под жизненным циклом понимают процесс репродукции вируса внутри инфицированной клетки. Особенности репродукции вирусов зависят от типа вирусного генома. Однако существуют некоторые общие закономерности
репродукции вирусов:
1.Все РНК-содержащие вирусы, кроме вирусов гриппа и ретровирусов,
репродуцируются в цитоплазме клетки. Геномы вирусов гриппа и ретровирусов при репродукции проникают в ядро инфицированной клетки.
2.Репродукция всех ДНК-содержащих вирусов, кроме вирусов оспы, протекает как в ядре, так и в цитоплазме клетки. При этом в ядре происходит транскрипция и репликация вирусных нуклеиновых кислот, а в цитоплазме клетки – трансляция вирусных белков и сборка дочерних вирионов. Размножение вирусов
202
оспы происходит полностью в цитоплазме клетки.
3.Нуклеокапсидные белки вирусов синтезируются на свободных полирибосомах (не связанных с мембраной), а суперкапсидные белки - на рибосомах, ассоциированных с мембранами.
4.Белки некоторых вирусов после образования полипептидной молекулы
подвергаются протеолитическому процессингу (расщеплению или разрезанию).
5.Суперкапсидные белки оболочечных вирусов при транспортировке к клеточной мембране подвергаются гликозилированию.
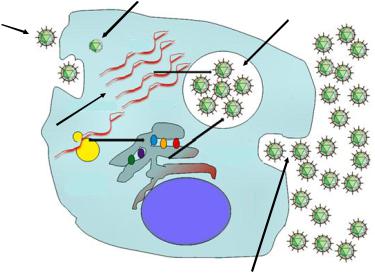
Жизненный цикл вирусов включает следующие этапы (рисунок 6.26):
1.Адсорбция вирусов на мембране клетки.
2.Проникновение вируса в клетку.
3.Депротеинизация (освобождение вирусной нуклеиновой кислоты).
4.Синтез компонентов вирусов (вирусных нуклеиновых кислот и вирусных структурных белков).
5.Формирование (сборка) зрелых дочерних вирионов.
6.Выход дочерних вирионов из клетки.
Депротеинизация
Адсорбция |
Сборка вирионов |
|
Проникновение 
Синтез вирусных компонентов 
Выход вирионов
Рисунок 6.26 – Этапы жизненного цикла вирусов. Заимствовано из Интернетресурсов.
Адсорбция вируса на мембране клетки происходит путем взаимодействия вирусного белка (антирецептора) с клеточными рецепторами. Для каждого вида вируса на клеточной мембране существуют специфические рецепторы, с которыми вирусы связываются. По химической природе рецепторы, на которых фиксируются вирусы, могут быть мукопротеиновыми или липопротеиновыми. За распознавание клеточных рецепторов отвечают белки вириона (капсидные или суперкапсидные).
Антирецепторы вирионов называются прикрепительными белками. Они имеют форму нитей, шипов, грибовидных структур. В процессе адсорбции важную роль играют электрические заряды: вирусы несут отрицательный заряд, а участки клеточной стенки - положительный заряд. Процесс адсорбции протекает в течение

203
5-90 минут. На одной клетке адсорбируется множество вирусных частиц, так как количество специфических рецепторов на поверхности одной клетки составляет 104- 105. Процесс адсорбции вируса на поверхности клетки представлен на рисунке 6.27.
Вирион
Прикрепительный
белок
Клеточный
рецептор
Клеточная мембрана
Рисунок 6.27 - Процесс адсорбции вируса на поверхности клетки.
Проникновение вируса в клетку происходит путем слияния вирусной оболочки с клеточной мембраной или путем рецептор-опосредованного эндоцитоза.
Путем слияния суперкапсида с клеточной мембраной в клетку проникают в основном оболочечные вирусы. В этом процессе участвуют специфические белки слияния. При этом происходит высвобождение нуклеокапсида в цитоплазму клетки
(рисунок 6.28).
Вирус
Клетка
 Ядро
Ядро
Рисунок 6.28 - Схема проникновения вируса в клетку путем слияния вирусной оболочки с клеточной мембраной.
Путем рецептор-опосредованного эндоцитоза в клетку проникают в основном безоболочечные вирусы. Вначале вирус связывается со специфическими рецепторами на клеточной поверхности. Затем происходит инвагинация (впячивание) клеточной мембраны, образование внутриклеточных вакуолей (эндосом) и их слияние с лизосомами. В дальнейшем вирусный геном в цитоплазме клетки высвобождается из эндосомы (рисунок 6.29).
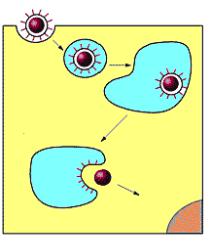
204
Вирус 
 Эндосома
Эндосома
Клетка 
 Ядро
Ядро
Рисунок 6.29 – Схема проникновения вируса путем рецепторопосредованного эндоцитоза.
Депротеинизация (освобождение вирусной нуклеиновой кислоты или раздевание вируса) происходит с помощью протеолитических ферментов клетки (протеаз и липаз). Смысл раздевания заключается в удалении вирусных оболочек. Конечными продуктами раздевания вирусов являются сердцевины, нуклеокапсиды или нуклеиновые кислоты. Для некоторых вирусов конечным продуктом являются нуклеиновые кислоты, связанные с внутренним вирусным белком (в частности, ферментом).
После депротеинизации вирусы невозможно выделить из культуры клеток. Этот этап репродукции известен как теневая фаза (фаза эклипса или затмения), во время которой вирус перестает существовать как оформленный вирион.
Синтез компонентов вирусов включает репликацию вирусных нуклеиновых кислот и синтез вирусных белков. В зависимости от типа вирусного генома (ДНК-содержащие или РНК-содержащие вирусы) синтез компонентов дочерних вирионов протекает либо в цитоплазме, либо в цитоплазме и ядре клетки.
У ДНК-содержащих вирусов проникший в цитоплазму нуклеокапсид транспортируется к ядру клетки. Вирусная ДНК проникает в клеточное ядро, где протекает транскрипция - переписывание информации с ДНК на РНК с помощью клеточной полимеразы. Исключение составляет ДНК-содержащий вирус оспы, у которого транскрипция протекает в цитоплазме клетки при участии вирусспецифического фермента (ДНК-полимеразы), который проникает в клетку в составе вириона. В результате транскрипции на матрице одной из нитей ДНК синтезируется иРНК, которая перемещается в цитоплазму клетки и запускает процесс трансляции – перевода генетической информации с иРНК на последовательность аминокислот в вирусных белках. Синтез белков происходит на рибосомах клетки. Одновременно в ядре клетки протекает процесс репликации (образование дочерних нуклеиновых кислот на матрице материнской ДНК). Синтезированные дочерние молекулы ДНК в составе нуклеокапсида перемещаются из ядра клетки в цитоплазму путем почкования, захватывая фрагмент ядерной мембраны. В цитоплазме клетки завершается сборка дочерних вирионов и их выход из клетки.
Таким образом, реализация генетической информации у ДНК-содержащих
205
вирусов идет следующим образом:
ДНК → транскрипция → иРНК → трансляция → белок
Плюс-РНК-содержащие вирусы адсорбируются на специфических клеточных рецепторах и проникают внутрь клетки путем эндоцитоза. Репликация плюс-РНК вирусов происходит в цитоплазме клеток. При этом функции иРНК выполняет вирусная нуклеиновая кислота. В результате трансляции на рибосомах образуется белковая молекула, которая разрезается клеточными протеазами на структурные и неструктурные вирусные белки. При этом образуется также полимераза, которая способствует образованию минус-РНК на матрице родительской плюс-РНК. На матрице минус-РНК происходит синтез молекул плюсРНК, участвующих в биосинтезе белков дочерних вирионов. Высвобождение дочерних вирионов сопровождается лизисом клетки или происходит путем почкования. У ретровирусов при репродукции образуется промежуточный продукт в виде молекулы ДНК.
Таким образом, реализация генетической информации у позитивных РНКсодержащих вирусов идет без этапа транскрипции:
плюс-РНК → трансляция → белок
Минус-РНК-содержащие вирусы проникают в клетку путем адсорбции или слияния с клеточной мембраной. Геном таких вирусов не может выполнять функцию иРНК, поэтому в цитоплазме клетки на матрице минус-РНК вначале синтезируется плюс-РНК. Этот процесс катализируется полимеразой (транскриптазой), находящейся в составе проникшего в клетку вириона. При синтезе плюс-РНК образуются полные нити и короткие нити. Короткие плюс-РНК- нити участвуют в синтезе ферментов и белков для дочерних популяций. Полные нити плюс-РНК служат матрицей для синтеза молекул минус-РНК дочерних вирионов. Дочерние вирионы транспортируются к модифицированным участкам клеточной мембраны и высвобождаются почкованием, захватывая фрагмент клеточной мембраны. Этот фрагмент клеточной мембраны служит для вирусной частицы суперкапсидом.
Таким образом, реализация генетической информации у минус-РНК- содержащих вирусов протекает по следующей схеме:
минус-РНК → транскрипция → иРНК → трансляция → белок
РНК-содержащие ретровирусы имеют уникальный путь репродукции. После проникновения в клетку генетическая информация с РНК этих вирусов переписывается на ДНК. Этот процесс называется обратной транскрипцией. Для его осуществления требуется специфический фермент - обратная транскриптаза или ревертаза. Этот фермент приносится в клетку в составе ретровирусов. Затем вновь образованная ДНК интегрирует с клеточным геномом и в его составе участвует в образовании иРНК, необходимой для синтеза вирусных белков. Транскрипцию интегрированной ДНК в составе клеточных геномов (переписывание
206
информации с ДНК на иРНК) осуществляет клеточная ДНК-зависимая РНКполимераза.
Таким образом, у ретровирусов отмечается следующий путь реализации генетической информации:
РНК → обратная транскрипция → ДНК → транскрипция → иРНК → трансляция → белок
В эукариотической клетке многие вирусные белки подвергаются
посттрансляционным модификациям: гликозилированию, ацилированию,
сульфированию, протеолитическому нарезанию и фосфорилированию. Гликозилирование белков заключается в присоединении к полипептиду углеводных остатков. Образовавшиеся гликопротеины входят в состав вирусных оболочек и находятся на поверхности вирусных частиц. Ацилирование белков заключается в присоединении к протеину 1-2 молекул жирных кислот. Сульфирование белков протекает путем связывания с углеводными остатками гликопротеина сульфатной группы. Протеолитическое нарезание характерно для многих вирусных белков. Оно осуществляется клеточными ферментами в специфических точках полипептидной молекулы. После этого белки приобретают функциональную активность. Фосфорилирование характерно для белков, связанных с вирусным геномом. Оно осуществляется как вирусными, так и клеточными ферментами.
Таким образом, синтез нуклеиновых кислот и вирусных белков протекает в разных структурах клетки, то есть синтез компонентов вирионов в клетке разобщен. Такой способ репродукции вирусов называется дизъюнктивным (разобщенным). Сборка дочерних вирионов возможна только при специфическом узнавании вирусных нуклеиновых кислот и белков и самопроизвольном их соединении друг с другом. У просто устроенных вирусов нуклеиновая кислота и белки взаимодействуют на мембранах эндоплазматического ретикулума, в результате чего формируется упорядоченная структура. У сложно устроенных вирусов сборка осуществляется многоступенчато. Вначале нуклеиновые кислоты взаимодействуют с внутренними белками, образуя нуклеокапсиды (сердцевины). Затем нуклеокапсиды выстраиваются с внутренней стороны клеточной мембраны под теми участками, которые модифицированы оболочечными вирусными белками. В результате этого происходит самосборка вирусных частиц. Количество зрелых вирионов, сформировавшихся в одной клетке, колеблется от 10 до 10000 и более.
Выход вирионов из клетки (высвобождение дочерних вирионов)
осуществляется двумя способами: путем лизиса клетки (взрывной способ выхода) и
путем почкования. Выход из клетки путем лизиса (“взрыва”) связан с деструкцией клетки. Такой способ характерен для безоболочечных вирусов, у которых отсутствует суперкапсидная оболочка. Выход вирионов из клетки путем почкования в основном характерен для оболочечных вирусов. При этом способе клетка может некоторое время сохранять жизнеспособность. Вирусы, содержащие суперкапсид, высвобождаются медленно (в течение 2-6 часов). Вначале у них суперкапсидные белки устанавливаются на наружной поверхности клеточной мембраны в виде своеобразных шипов, вытесняя клеточные белки. Затем через

207
модифицированную клеточную мембрану проходит нуклеокапсид с образованием суперкапсидной оболочки. Весь цикл репродукции у РНК-вирусов продолжается 4-8 часов, а у ДНК-вирусов – 12-24 часа.
6.9. Культивирование вирусов
Культивирование вирусов проводят при изучении клиники и патогенеза вирусных заболеваний, при диагностике вирусных инфекций, а также для приготовления диагностических и вакцинных препаратов, используемых в вирусологии. Культивирование вирусов осуществляют в трех биологических системах:
-в организме лабораторных животных;
-в развивающихся куриных эмбрионах;
-в культурах клеток.
Для заражения биологических систем из исследуемого материала готовят суспензию, в которую добавляют антибиотики для освобождения от посторонней микрофлоры (бактерий и грибов). После этого исследуемый материал вводят в
биологическую систему.
Лабораторные животные. В вирусологических исследованиях используют белых мышей, белых крыс, морских свинок, золотистых хомячков, кроликов и других лабораторных животных (рисунок 6.31).
а б Рисунок 6.31 – Лабораторные животные, используемые в вирусологических
исследованиях: а – белая мышь, б – золотистый хомячок. Заимствовано из Интернет-ресурсов.
Лабораторных животных заражают вируссодержащим материалом различными способами (подкожно, внутримышечно, внутривенно, интраназально, интрацеребрально и т. д.). Способ заражения выбирают в зависимости от тропизма (избирательности поражения тканей) вирусов (рисунок 6.32).

208
а б Рисунок 6.32 – Внутрибрюшинное (а) и интрацеребральное (б) заражение
белых мышей. Заимствовано из Интернет-ресурсов.
О репродукции вирусов в организме животных судят по развитию у них видимых клинических проявлений заболевания, патоморфологическим изменениям органов и тканей, а также на основании реакции гемагглютинации (РГА) с суспензией из органов, содержащих вирусы. РГА основана на способности многих вирусов вызывать склеивание (агглютинацию) эритроцитов человека, птиц и млекопитающих в результате взаимодействия вирусных белков (гемагглютининов) с рецепторами эритроцитов.
Развивающиеся куриные эмбрионы (РКЭ) в вирусологических исследованиях используют в возрасте 5-12 дней. С использованием развивающихся куриных эмбрионов возможно приготовить большое количество вируссодержащего материала, что особенно важно в производстве вакцин или других лечебнопрофилактических препаратов.
Способы заражения РКЭ (рисунок 6.33):
-на ХАО;
-в аллантоисную полость;
-в амниотическую полость;
-в желточный мешок;
-в тело эмбриона.
Втело эмбриона
|
Хорионаллантоисная оболочка |
Аллантоисная полость |
Амниотическая полость |
Валлантоисную полость
Вжелточный мешок
Желточный мешок
Рисунок 6.33 – Способы заражения куриного эмбриона. Заимствовано из Интернет-ресурсов.

209
Признаками репродукции вирусов в РКЭ является гибель эмбриона, патологоанатомические изменения в оболочках и тканях эмбриона (оспины, кровоизлияния), положительная реакция гемагглютинации. О гибели свидетельствует прекращение движений эмбриона, выявляемое при их овоскопировании, то есть просвечивании (рисунок 6.34).
Рисунок 6.34 – Овоскопирование куриных яиц. Заимствовано из Интернетресурсов.
Погибшие эмбрионы вскрывают, содержимое извлекают (рисунок 6.35) и проводят патологоанатомическое исследование. Эмбриональную жидкость смешивают с эритроцитами для определения гемагглютинации.
Рисунок 6.35 – Вскрытие куриного эмбриона. Заимствовано из Интернетресурсов.
Наиболее широкое распространение для культивирования вирусов получила культура клеток. Клеточную культуру получают из органов и тканей человека, животных, птиц. Метод получения культуры клеток для выращивания вирусов разработали американские вирусологи Дж. Эндерс (John Franklin Enders, 1897-1985
гг.), Т. Уэллер (Thomas Huckle Weller, 1915-2008 гг.) и Ф. Роббинс (Frederick Chapman Robbins, 1916-2003 гг.). В 1954 г. они получили Нобелевскую премию за технику культивирования вируса полиомиелита в тканевых культурах. Клетки, полученные из различных органов и тканей, размножают вне организма на искусственных питательных средах в специальной лабораторной посуде (рисунок
6.36).

210
Рисунок 6.36 – Посуда для клеточных культур.
При выращивании клеточных культур необходимо выполнять следующие условия:
-соблюдать правила асептики;
-использовать лабораторную посуду из нейтрального стекла;
-использовать сложные по составу питательных сред (среда 199, Игла), содержащие минеральные соли, аминокислоты, витамины, глюкозу, сыворотку крови животных или человека, буферные растворы;
-добавлять к питательной среде антибиотики для подавления роста посторонних микробов;
-соблюдать оптимальную температуру культивирования (36-38,5ОС).
По способу культивирования клеточные культуры подразделяются на следующие виды:
1.Однослойные (монослойные) культуры клеток – клетки способны прикрепляться и размножаться на поверхности химически нейтрального стекла лабораторной посуды в виде монослоя.
2.Суспензионные культуры клеток – клетки способны размножаться во всем объеме питательной среды при постоянном ее перемешивании. Модификацией этого метода является проточное культивирование, при котором в специальный аппарат непрерывно добавляют свежую питательную среду и удаляют отработанную.
3.Смешанные культуры клеток – способны культивироваться как на поверхности подложки, так и в суспензии.
По способу получения культуры клеток подразделяются на следующие
виды:
1.Органные культуры – цельные кусочки органов и тканей, сохраняющие исходную структуру вне организма.
2.Первичные культуры клеток способны размножаться только в первых генерациях, то есть выдерживают не более 5-10 пассажей после выделения из тканей. Первичные культуры получают путем обработки кусочков тканей (нормальной, опухолевой, эмбриональной) протеолитическими ферментами (трипсином, коллагеназой), которые разрушают межклеточные связи в тканях с образованием изолированных клеток. Разобщенные клетки помещают в питательную среду и вносят в стеклянный сосуд. Клетки прикрепляются к стенке сосуда и размножаются, образуя монослой (слой толщиной в одну клетку). С помощью специальных веществ (трипсина, версена) клетки можно снять с поверхности сосуда и перенести в другой. Такая манипуляция называется пассажем.