

Соперничество между семью субъединицами за кор-фермент
Для
сравнения сродства связывания
кор-ферментов между семью σ субъединицами
E. coli мы очистили все остальные шесть
субъединиц σ от сверхэкспрессированных
клеток E. coli до кажущейся однородности
(рис. 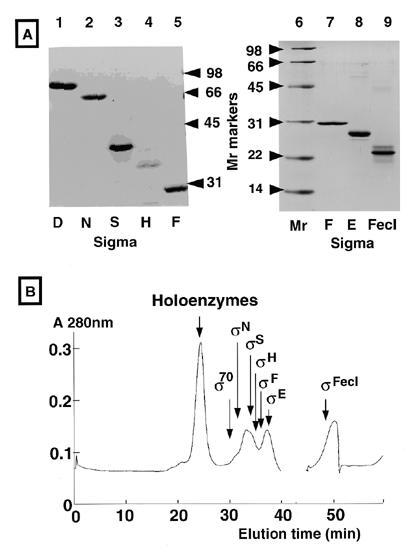 2А).
Чтобы восстановить все семь субъединиц
σ в активных формах, была предпринята
попытка очистить σ субъединицы той же
самой процедурой от сверхэкспрессированных
клеточных экстрактов, т. е. извлечение
σ белков из фракции гранул
сверхэкспрессированных клеточных
экстрактов с использованием буфера
солюбилизации, содержащего неионный
детергент, без использования ионных
детергентов. Концентрацию каждой
субъединицы σ определяли после коррекции
на чистоту (90-98%), согласно оценке SDS-PAGE
с последующим окрашиванием CBB. Так как
все эти очищенные субъединицы σ полностью
превращались в соответствующие
холоферменты путем добавления избыточных
количеств кор-фермента (данные не
показаны), используемые препараты σ
были полностью активны, по крайней мере,
в связывании с кор-ферментом.
2А).
Чтобы восстановить все семь субъединиц
σ в активных формах, была предпринята
попытка очистить σ субъединицы той же
самой процедурой от сверхэкспрессированных
клеточных экстрактов, т. е. извлечение
σ белков из фракции гранул
сверхэкспрессированных клеточных
экстрактов с использованием буфера
солюбилизации, содержащего неионный
детергент, без использования ионных
детергентов. Концентрацию каждой
субъединицы σ определяли после коррекции
на чистоту (90-98%), согласно оценке SDS-PAGE
с последующим окрашиванием CBB. Так как
все эти очищенные субъединицы σ полностью
превращались в соответствующие
холоферменты путем добавления избыточных
количеств кор-фермента (данные не
показаны), используемые препараты σ
были полностью активны, по крайней мере,
в связывании с кор-ферментом.
Используя очищенные субъединицы σ, мы сначала проанализировали кривую насыщения каждой субъединицы σ, чтобы преобразовать фиксированное количество (20 пмоль) кор-фермента в соответствующий холофермент. При входном молярном соотношении 1 более 80% входного кор-фермента было преобразовано в холофермент для пяти субъединиц σ: σ70, σN, σF, σE и σFecI, а 60-65% входного кор-фермента было преобразовано в холофермент для σS и σH (данные не показаны). Для насыщения σ входного кор-фермента требовалось более высокой концентрации как для σS, так и для σH. Входной кор-фермент насыщали добавлением 2-кратного молярного избытка σ даже для σS и σH, со слабым сродством связывания. Аналогичный порядок активности σ был получен при определении уровня функционального холофермента путем измерения кривой насыщения σ с использованием транскрипционных тестов in vitro, направленных специфическими промоторами для каждой субъединицы σ (данные не показаны). Однако при наличии единичных σ-добавок было трудно определить небольшое различие в сродстве связывания кор-фермента между семью σ-субъединицами.
Д ля
того чтобы измерить относительное
сродство связывания кор-фермента между
семью субъединицами σ, мы провели
смешанный эксперимент восстановления
в присутствии всех семи субъединиц σ в
одной и той же реакционной смеси. К 20
пмоль кор-фермента добавляли увеличивающиеся
количества эквимолярной смеси всех
семи субъединиц σ, а субъединицы σ
фракционировали на связанные с
кор-ферментом и несвязанные фракции
методом гель-фильтрационной колоночной
хроматографии (рис. 2B). После измерения
молярных соотношений между α-субъединицей
и каждой σ-субъединицей в пике холофермента
определяли основные связанные с ферментом
σ-субъединицы. Почти 68% кор-фермента
было преобразовано в холоферменты при
входном молярном отношении 0,25 для каждой
субъединицы σ (или 1,75 для комбинированных
субъединиц σ) (рис. 3А) и >90% входного
кор-фермента связывалось с одной из
семи субъединиц σ при входном молярном
отношении 1,0 для каждой субъединицы σ
(или 7,0 для комбинированных субъединиц
σ) (рис. 3Б). Входной фермент ядра насыщался
одной из субъединиц σ при входном
молярном отношении 2,0 для каждой
субъединицы σ (или 14 для комбинированных
субъединиц σ) (рис. 3С). Однако
кор-фермент-связанная фракция существенно
отличалась среди семи субъединиц σ. При
низкой концентрации σ (или уровне
насыщения 60-70%) σN
проявлял сродство столь же высокое, как
и субъединица σ70,
а комбинированный уровень EσN
и Eσ70
достигал 60% от общего количества
холоферментов. Уровень холоферментов
был почти одинаковым среди EσF,
EσFecI
и
EσE,
каждый из которых составлял 14, 10 и 8%
соответственно, в то время как количество
EσH
составлял <1% от общего количества
холоферментов. Уровень образования EσS
был самым низким при использованных
условиях восстановления. Фракции,
связанные с кор-ферментом, увеличивались
параллельно для всех семи субъединиц
σ одновременно с увеличением входных
субъединиц σ до фиксированного количества
основного фермента (рис. 3B и C).
ля
того чтобы измерить относительное
сродство связывания кор-фермента между
семью субъединицами σ, мы провели
смешанный эксперимент восстановления
в присутствии всех семи субъединиц σ в
одной и той же реакционной смеси. К 20
пмоль кор-фермента добавляли увеличивающиеся
количества эквимолярной смеси всех
семи субъединиц σ, а субъединицы σ
фракционировали на связанные с
кор-ферментом и несвязанные фракции
методом гель-фильтрационной колоночной
хроматографии (рис. 2B). После измерения
молярных соотношений между α-субъединицей
и каждой σ-субъединицей в пике холофермента
определяли основные связанные с ферментом
σ-субъединицы. Почти 68% кор-фермента
было преобразовано в холоферменты при
входном молярном отношении 0,25 для каждой
субъединицы σ (или 1,75 для комбинированных
субъединиц σ) (рис. 3А) и >90% входного
кор-фермента связывалось с одной из
семи субъединиц σ при входном молярном
отношении 1,0 для каждой субъединицы σ
(или 7,0 для комбинированных субъединиц
σ) (рис. 3Б). Входной фермент ядра насыщался
одной из субъединиц σ при входном
молярном отношении 2,0 для каждой
субъединицы σ (или 14 для комбинированных
субъединиц σ) (рис. 3С). Однако
кор-фермент-связанная фракция существенно
отличалась среди семи субъединиц σ. При
низкой концентрации σ (или уровне
насыщения 60-70%) σN
проявлял сродство столь же высокое, как
и субъединица σ70,
а комбинированный уровень EσN
и Eσ70
достигал 60% от общего количества
холоферментов. Уровень холоферментов
был почти одинаковым среди EσF,
EσFecI
и
EσE,
каждый из которых составлял 14, 10 и 8%
соответственно, в то время как количество
EσH
составлял <1% от общего количества
холоферментов. Уровень образования EσS
был самым низким при использованных
условиях восстановления. Фракции,
связанные с кор-ферментом, увеличивались
параллельно для всех семи субъединиц
σ одновременно с увеличением входных
субъединиц σ до фиксированного количества
основного фермента (рис. 3B и C).
При насыщении порядок кор-фермент-связанных субъединиц σ составлял σ70 > σN > σF > σH/σFecI > σE > σS. В условиях насыщения σ доля холофермента Eσ70 составляла ∼39% от общего количества холоферментов. Отсюда мы заключаем, что в условиях конкурентного связывания в присутствии всех семи субъединиц σ сродство σ70 является самым сильным, а сродство σS-самым слабым. Судя по разнице сродства (>16 раз) в связывании кор-фермента между σ70 и σS и значению Kd = 0,26 нм для σ70 (см. выше), мы оцениваем, что кажущееся значение KD для σS с кор-ферментом составляет ≈ 4,3 нм (табл.1).
-
σ subunit
Kd (nM)a
Core binding affinity
Intracellular concentrationb
Holoenzyme ratioc (%)
(fmol/mg protein)
(molecules/cell)
σ70 (σD)
0.26
1.0
160
700
78
σ54 (σN)
0.30
1.55
25
110
8
σ38 (σS)
4.26
16.4
<1
<1
0
σ32
(σH)
1.24
4.75
2.1
<10
0
σ28
(σF)
0.74
2.85
85
370
14
σ24
(σE)
2.43
9.35
1.4
<10
0
σ18
(σFecI)
1.73
6.65
<1
<1
0