

Strukov_A_I_Serov_V_V_Patanatomia_Uchebnik_20
.pdfагентами. «Бреши» в мембране, независимо от того, «сквозные» они или «поверхностные», ведут к осмотическому набуханию клетки и ее гибели.
«Штопка» локально разрушенной плазмолеммы осуществляется с помощью мембран мелких везикул, которые сосредоточиваются в месте повреждения.
Своеобразным изменением плазмолеммы, встречающимся не только при нарушении ее проницаемости, является образование миелиноподобных структур (рис. 24). Эти структуры появляются в связи с перекисным окислением липидов мембран, усиливающимся под воздействием разных агентов. Высвобождающиеся из разрушающихся при перекисном окислении мембран фосфолипиды (дезагрегация и реагрегация мембраны) образуют сложные миелиноподобные структуры. Подобные структуры появляются и при скручивании удлиненных цитоплазматических отростков.
Изменения клетки при повреждении плазмолеммы. Повреждение плазмолеммы ведет к утрате так называемого активного мембранного транспорта: концентрации интра- и экстрацеллюлярного натрия и калия выравниваются, внутрь клетки проникают низкомолекулярные анионы, а затем и катионы, повышается внутриклеточное осмотическое давление. Таким образом, резко нарушается мембранный водно-электролитный транспорт, следствием чего становятся набухание и отек клетки. Нарушение активного мембранного транспорта может приводить также к избирательному поступлению в клетку определенных продуктов обмена (белки, липиды, углеводы, пигменты) и накоплению их после истощения ферментных систем, метаболизирующих эти продукты. Так развиваются клеточные дистрофии инфильтрационного генеза(жировая дистрофия гепатоцитов при гиперлипидемиях; гиалиново-капельная дистрофия нефроцитов при нефротическом синдроме). При резком повреждении плазмолеммы и поступлении в клетку ряда токсических или биологически активных веществ возможна деструкция структурных комплексов клетки
51

Рис.
24. Миелиноподобные структуры под плазматической мембраной мышечного волокна при ишемии. х22 500
с высвобождением составляющих их химических веществ (белки, липиды и т.д.), что ведет к их накоплению. Возникают клеточные дистрофии декомпозиционного генеза (жировая дистрофия миокарда при дифтерии, гидропическая дистрофия гепатоцитов при вирусном гепатите). Следует заметить, что инфильтрационный механизм развития дистрофии может сменяться декомпозиционным и наоборот. В ряде случаев повреждения плазмолеммы позволяют проникнуть в клетку веществам, способным извратить синтез того или иного продукта. Тогда возникают клеточные дистрофии извращенного синтеза (синтез алкогольного гиалина гепатоцитом под воздействием этанола). Финалом тяжелого повреждения плазмолеммы является гибель клетки - ее некроз(см. Дистрофия, Некроз).
Патология клеточных стыков
В тканях человека клеточные стыки ответственны за три главные функции: межклеточную адгезию, «тесное общение» клеток и герметизацию слоя эпителиальных клеток.
Межклеточную адгезию как чисто механическую функцию ранее связывали в первую очередь с десмосомами. В настоящее время установлено, что в межклеточной адгезии участвуют все типы клеточных стыков.
Медиаторами «тесного общения» (или сопряжения) клеток считают щелевидные стыки,
которые обеспечивают прямое сообщение между клетками, перенос ионов и малых молекул без потери их во внеклеточное пространство. Это способствует регуляции метаболических процессов в клетках и их дифференцировке.
52
Герметизация клеток эпителиального пласта обеспечивается плотными стыками, степень ее коррелирует с количеством стыков и внутримембранных тяжей. Плотные стыки отвечают за поддержание осмотических и электрохимических градиентов эпителиального пласта и отчасти за состояние внеклеточных структур, окружающих этот пласт.
Изменение межклеточной адгезии. Показано, что степень межклеточной адгезии ослабевает при опухолевом росте, причем уже на ранних стадиях онкогенеза. Количество и распределение клеточных стыков на поверхности опухолевых клеток могут быть одним из критериев характеристики роста опухоли.
Изменение «тесного общения» клеток. Как уже говорилось, «тесное общение» клеток предопределяет их непосредственный контакт для обмена информационными молекулами и обычно осуществляется с помощью щелевидных стыков, гидрофильные каналы которых пропускают ионы и молекулы с молекулярной массой до 1000. Считают, что дефекты «тесного общения» клеток могут играть важную роль в развитии и поведении опухолей.
Нарушения межмембранных связей клеток тканевых барьеров. Плотные стыки являются структурной основой таких тканевых барьеров, как кровь - мозг, кровь - легкие, кровь - желчь, кровь - почки. Поэтому эти стыки находятся, как правило, в эпителии. Они предотвращают «про-
извольный обмен» белками и другими макромолекулами между клеточными «партнерами» барьеров. Наиболее частым следствием повреждения тканевых барьеров является увеличение проницаемости плотных стыков клеток (рис. 25), что ведет к «трансэпителиальной протечке» (например, при повышении внутрисосудистого гидростатического давления, мозговой коме, холестазе, шоке, нефротическом синдроме).
53

Рис. 25. Расхождение десмосомальных контактов между гепатоцитами (показано стрелками) вблизи желчного канальца при первичном билиарном циррозе. х23 500
Структурные изменения клеточных стыков. Эти изменения касаются прежде всего десмосом.Псевдодесмосомы («несовершенные» десмосомы) с хорошо развитой пластинкой лишь у одной клетки могут возникать в результате разрыва дефектных стыков, неполной сборки стыка, диссоциации клеток. В основеасимметричных десмосом с недоразвитой пластинкой у одной из клеток лежат, вероятно, те же механизмы. К структурным изменениям клеточных стыков следует отнести и нарушения их топографии, т.е. появление их на поверхности клеток, где они в обычных условиях жизнедеятельности клеток не встречаются.
Изменения структуры десмосом, как и других типов клеточных стыков, находят при метаплазии, дисплазии, опухолевом росте, в эмбриональных тканях (асимметричные десмосомы); они найдены при таких заболеваниях, как ревматоидный артрит, псориаз.
В заключение следует сказать, что патология клетки как интегративное понятие является необходимой базой общей патологии человека.
ДИСТРОФИЯ
Общие сведения
54
Дистрофия (от греч. dys - нарушение и trophe - питаю) - сложный патологический процесс, в основе которого лежит нарушение тканевого (клеточного) метаболизма, ведущее к структурным изменениям. Поэтому дистрофии рассматриваются как один из видов повреждения.
Под трофикой понимают совокупность механизмов, определяющих метаболизм и структурную организацию ткани (клетки), которые необходимы для отправления специализированной функции. Среди этих механизмов выделяют клеточные и внеклеточные (рис. 26). Клеточные механизмы обеспечиваются структурной организацией клетки и ее ауторегуляцией. Это значит, что трофика клетки в значительной мере явля-
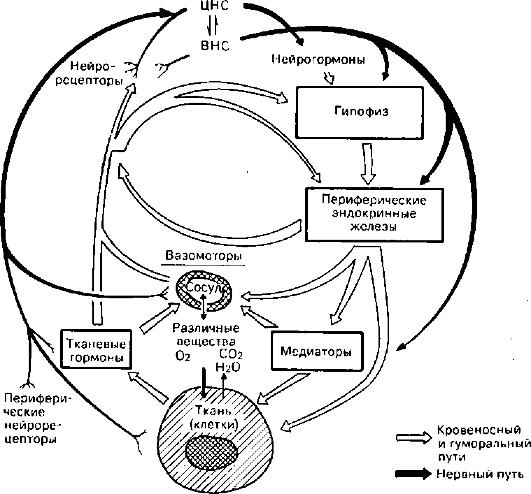 Рис.
Рис.
26. Механизмы регуляции трофики (по М.Г. Балш)
ется свойством самой клетки как сложной саморегулирующейся системы. Жизнедеятельность клетки обеспечивается «окружающей средой» и регулируется с помощью ряда систем организма. Поэтому внеклеточные механизмы трофики располагают транспортными (кровь, лимфа, микроциркуляторное русло) и интегративными (нейро-эндокринные, нейрогуморальные) системами ее регуляции. Из сказанного следует, чтонепосредственной причиной развития дистрофий могут служить нарушения как клеточных, так и внеклеточных механизмов, обеспечивающих трофику.
55
1. Расстройства ауторегуляции клетки могут быть вызваны различными факторами (гиперфункция, токсические вещества, радиация, наследственная недостаточность или отсутствие фермента и т.д.). Большую роль придают полому генов - рецепторов, осуществляющих «координированное торможение» функций различных ультраструктур.
Нарушение ауторегуляции клетки ведет к энергетическому ее дефициту и к нарушению ферментативных процессов в клетке. Ферментопатия, или энзимопатия (приобретенная или наследственная), становится основным патогенетическим звеном и выражением дистрофии при нарушениях клеточных механизмов трофики.
2.Нарушения функции транспортных систем, обеспечивающих метаболизм и структурную сохранность тканей (клеток), вызывают гипоксию, которая является ведущей в патогенезе дисциркуляторных дистрофий.
3.При расстройствах эндокринной регуляции трофики (тиреотоксикоз, диабет, гиперпаратиреоз и т.д.) можно говорить об эндокринных, а при нарушении нервной регуляции трофики (нарушенная иннервация, опухоль головного мозга и т.д.) - о
нервных или церебральных дистрофиях.
Особенности патогенеза внутриутробных дистрофий определяются непосредственной связью их с болезнями матери. В исходе при гибели части зачатка органа или ткани может развиться необратимый порок развития.
При дистрофиях в клетке и (или) межклеточном веществе накапливаются различные продукты обмена (белки, жиры, углеводы, минералы, вода), которые характеризуются количественными или качественными изменениями в результате нарушения ферментативных процессов.
Морфогенез. Среди механизмов, ведущих к развитию характерных для дистрофий изменений, различают инфильтрацию, декомпозицию (фанероз), извращенный синтез и трансформацию.
Инфильтрация - избыточное проникновение продуктов обмена из крови и лимфы в клетки или межклеточное вещество с последующим их накоплением в связи с недостаточностью ферментных систем, метаболизирующих эти продукты. Таковы, например, инфильтрация грубодисперсным белком эпителия проксимальных канальцев почек при нефротическом синдроме, инфильтрация холестерином и липопротеидами интимы аорты и крупных артерии при атеросклерозе.
Декомпозиция (фанероз) - распад ультраструктур клеток и межклеточного вещества, ведущий к нарушению тканевого (клеточного) метаболизма и накоплению продуктов нарушенного обмена в ткани (клетке). Таковы жи-
ровая дистрофия кардиомиоцитов при дифтерийной интоксикации, фибриноидное набухание соединительной ткани при ревматических болезнях.
56
Извращенный синтез - это синтез в клетках или в тканях веществ, не встречающихся в них в норме. К ним относятся: синтез аномального белка амилоида в клетке и аномальных белковополисахаридных комплексов амилоида в межклеточном веществе; синтез белка алкогольного гиалина гепатоцитом; синтез гликогена в эпителии узкого сегмента нефрона при сахарном диабете.
Трансформация - образование продуктов одного вида обмена из общих исходных продуктов, которые идут на построение белков, жиров и углеводов. Такова, например, трансформация компонентов жиров и углеводов в белки, усиленная полимеризация глюкозы в гликоген и др.
Инфильтрация и декомпозиция - ведущие морфогенетические механизмы дистрофий - часто являются последовательными стадиями в их развитии. Однако в некоторых органах и тканях в связи со структурнофункциональными их особенностями преобладает какой-либо один из морфогенетических механизмов (инфильтрация - в эпителии почечных канальцев, декомпозиция - в клетках миокарда), что позволяет говорить об ортологии (от греч. orthos - прямой, типичный) дистрофий.
Морфологическая специфика. При изучении дистрофий на разных уровнях - ультраструктурном, клеточном, тканевом, органном - морфологическая специфика проявляется неоднозначно. Ультраструктурная морфология дистрофий обычно не имеет какой-либо специфики. Она отражает не только повреждение органелл, но и их репарацию (внутриклеточная регенерация). Вместе с тем возможность выявления в органеллах ряда продуктов обмена (липиды, гликоген, ферритин) позволяет говорить об ультраструктурных изменениях, характерных для того или иного вида дистрофий.
Характерная морфология дистрофий выявляется, как правило, на тканевом и клеточном уровнях, причем для доказательства связи дистрофии с нарушениями того или иного вида обмена требуется применение гистохимических методов. Без установления качества продукта нарушенного обмена нельзя верифицировать тканевую дистрофию, т.е. отнести ее к белковым, жировым, углеводным или другим дистрофиям. Изменения органа при дистрофии (размер, цвет, консистенция, структура на разрезе) в одних случаях представлены исключительно ярко, в других - отсутствуют, и лишь микроскопическое исследование позволяет выявить их специфичность. В ряде случаев можно говорить о системном характере изменений при дистрофии (системный гемосидероз, системный мезенхимальный амилоидоз, системный липоидоз).
В классификации дистрофий придерживаются нескольких принципов. Выделяют дистрофии.
I. В зависимости от преобладания морфологических изменений в специализированных элементах паренхимы или строме и сосудах: 1) паренхиматозные; 2) стромально-сосудистые; 3) смешанные.
II. По преобладанию нарушений того или иного вида обмена: 1) белковые; 2) жировые; 3) углеводные; 4) минеральные.
57
III. В зависимости от влияния генетических факторов: 1) приобретенные; 2) наследственные.
IV. По распространенности процесса: 1) общие; 2) местные.
Паренхиматозные дистрофии
Паренхиматозные дистрофии - проявления нарушений обмена в высокоспециализированных в функциональном отношении клетках. Поэтому при паренхиматозных дистрофиях преобладают нарушения клеточных механизмов трофики. Различные виды паренхиматозных дистрофий отражают недостаточность определенного физиологического (ферментативного) механизма, служащего выполнению специализированной функции клеткой (гепатоцит, нефроцит, кардиомиоцит и т.д.). В связи с этим в разных органах (печень, почки, сердце и т.д.) при развитии одного и того же вида дистрофии участвуют различные пато- и морфогенетические механизмы. Из этого следует, что переход одного вида паренхиматозной дистрофии в другой вид исключается, возможно лишь сочетание разных видов этой дистрофии.
В зависимости от нарушений того или иного вида обмена паренхиматозные дистрофии делят на белковые (диспротеинозы), жировые (липидозы) и углеводные.
Паренхиматозные белковые дистрофии (диспротеинозы)
Большая часть белков цитоплазмы (простых и сложных) находится в соединении с липидами, образуя липопротеидные комплексы. Эти комплексы составляют основу мембран митохондрий, эндоплазматической сети, пластинчатого комплекса и других структур. Помимо связанных белков, в цитоплазме содержатся и свободные. Многие из последних обладают функцией ферментов.
Сущность паренхиматозных диспротеинозов состоит в изменении физико-химических и морфологических свойств белков клетки: они подвергаются денатурации и коагуляции или, наоборот, колликвации, что ведет к гидратации цитоплазмы; в тех случаях, когда нарушаются связи белков с липидами, возникает деструкция мембранных структур клетки. В исходе этих нарушений может развиться коагуляционный (сухой)
иликолликвационный (влажный) некроз (схема I).
Кпаренхиматозным диспротеинозам относят гиалиново-капельную, гидропическую и роговую дистрофии.
Кпаренхиматозным белковым дистрофиям со времен Р. Вирхова причисляли и многие патологи продолжают причислять так называемую зернистую дистрофию, при которой в клетках паренхиматозных органов появляются белковые зерна. Сами органы увеличиваются в размерах, становятся дряблыми и тусклыми на разрезе, что послужило причиной называть также зернистую дистрофию тусклым (мутным) набуханием.Однако электронномикроскопическое и гистоферменто-
Схема I. Морфогенез паренхиматозных диспротеинозов
58

химическое изучение «зернистой дистрофии» показало, что в ее основе лежит не накопление белка в цитоплазме, а гиперплазия ультраструктур клеток паренхиматозных органов как выражение функционального напряжения этих органов в ответ на различные воздействия; гиперплазированные ультраструктуры клетки выявляются при светооптическом исследовании как белковые гранулы.
Гиалиново-капельная дистрофия
При гиалиново-капельной дистрофии в цитоплазме появляются крупные гиалиноподобные белковые капли, сливающиеся между собой и заполняющие тело клетки; при этом происходит деструкция ультраструктурных элементов клетки. В ряде случаев гиалиново-капельная дистрофия завершается фокальным коагуляционным некрозом клетки.
Этот вид диспротеиноза часто встречается в почках, редко - в печени и совсем редко - в миокарде.
В почках при микроскопическом исследовании накопление гиалиновых капель находят в нефроцитах. При этом наблюдается деструкция митохондрий, эндоплазматической сети, щеточной каемки (рис. 27). В основе гиалиново-капельной дистрофии нефроцитов лежит недостаточность вакуолярно-лизосомального аппарата эпителия проксимальных канальцев, в норме реабсорбирующего белки. Поэтому этот вид дистрофии нефроцитов очень часто встречается при нефротическом синдроме. Этот синдром является одним из проявлений многих заболеваний почек, при которых первично поражается гломерулярный фильтр (гломерулонефрит, амилоидоз почек, парапротеинемическая нефропатия и др.).
Внешний вид почек при этой дистрофии не имеет каких-либо характерных черт, он определяется прежде всего особенностями основного заболевания (гломерулонефрит, амилоидоз).
В печени при микроскопическом исследовании в гепатоцитах находят гиалиноподобные тельца (тельца Мэллори), которые состоят из фибрилл
59
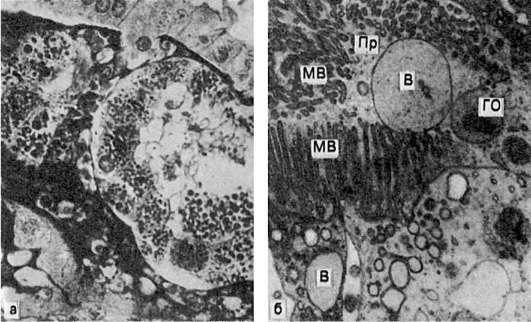
Рис.
27. Гиалиново-капельная дистрофия эпителия почечных канальцев:
а - в цитоплазме эпителия крупные белковые капли (микроскопическая картина); б - в цитоплазме клетки много белковых (гиалиновых) образований (ГО) овальной формы и вакуолей (В); отмечаются десквамация микроворсинок (МВ) щеточной каемки и выход в просвет (Пр) канальца вакуолей и белковых образований. Электронограмма. х18 000
особого белка - алкогольного гиалина (см. рис. 22). Образование этого белка и телец Мэллори служит проявлением извращенной белковосинтетической функции гепатоцита, что встречается постоянно при алкогольном гепатите и сравнительно редко при первичном билиарном и индийском детском циррозах, гепатоцеребральной дистрофии (болезни Вильсона-Коновалова).
Внешний вид печени различен; изменения характерны для тех ее заболеваний, при которых встречается гиалиново-капельная дистрофия.
Исход гиалиново-капельной дистрофии неблагоприятен: она завершается необратимым процессом, ведущим к некрозу клетки.
Функциональное значение этой дистрофии очень велико. С гиалиновокапельной дистрофией эпителия почечных канальцев связаны появление в моче белка (протеинурия) и цилиндров (цилиндрурия), потеря белков плазмы (гипопротеинемия), нарушение ее электролитного баланса. Гиалиново-капельная дистрофия гепатоцитов нередко является морфологической основой нарушений многих функций печени.
Гидропическая дистрофия
Гидропическая, или водяночная, дистрофия характеризуется появлением в клетке вакуолей, наполненных цитоплазматической жидкостью. Она наблюдается чаще в эпителии кожи и почечных канальцев, в гепа-
60