

Протисты / Карпов С.А. Строение клеток протистов
.pdfГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
Рис. 2.67. Внешний вид клеток Chlorarachnion. (Ïî: Van den Hoek et al., 1995.)
А – часть плазмодия с тремя вегетативными клетками и ветвяшимися ретикулоподиями (р), Б – одножгутиковая зооспора, В – амебоидная клетка, Г – коккоидная клетка с оболочкой. п – захваченный пищевой объект, пи – пиреноид, р – ретикулоподии, хл – хлоропласт, я – ядро.
Отряд Spongomonadida (Hibberd) Karpov, 1990 Спонгомонады (Ðèñ. 2.68)
Эти организмы обычно образуют прикрепленные колонии, клетки которых имеют 1 или чаще 2 жгутика, погружены в основу из слизи и железосодержащих гранул эндогенного происхождения. В митохондриях трубчатые или пузырьковидные
110
кристы. Основание жгутиков защищено цитоплазматическим выростом переднего конца клетки. Особенности питания и размножения не изучены. По данным молекулярной филогении, положение этих организмов в системе неопределенно: Phalansterium иногда группируется с апузомонадами, а Spongomonas – с хлорарахниевыми водорослями.
Представители: Spongomonas, Rhipidodendron, Phalansterium.
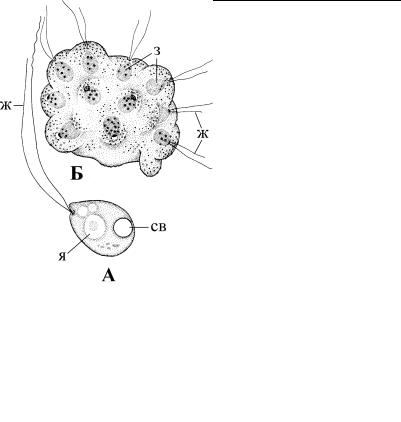
Рис. 2.68. Внешний вид спонгомонады Spongomonas. (По: Patterson, Hedley, 1992.) А – одиночная клетка, Б – колония. ж – жгутики, з – зооиды колонии, св –
сократительная вакуоль, я – ядро.
«Excavata» (Simpson) Экскаваты (Ðèñ. 2.69)
Небольшая группа свободноживущих протистов с 2–4 жгутиками, один из которых (как у ретортамонад) направлен назад, имеет характерные боковые выросты (кили) и проходит в вентральной бороздке, заканчивающейся цитофаринксом. Вентральная бороздка, как и у ретортамонад, укреплена развитыми корешками. Чаще всего встречаются в анаэробных условиях, многие не имеют митохондрий.
В настоящее время группа находится в стадии интенсивного изучения. По признакам ультратонкого строения экскаваты
111
ГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
сходны с ретортамонадами и могут быть отнесены к этому отряду. По данным молекулярной филогении, одни виды близки к ретортамонадам, другие – к гетеролобозным амебам.
Группа включает 6 родов: Jacoba, Malawimonas, Reclinimonas, Histiona, Trimastix è Carpediomonas.
Кроме рассмотренных таксонов, существует очень много протистов как с изученным, так и неизвестным ультратонким строением, положение которых в системе эукариот не опреде-
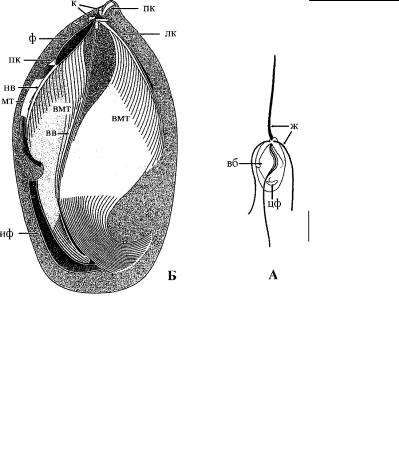
Рис. 2.69. Общий вид (А) и схема строения вентрального скелета клетки (Б) Trimastix marina. (По: Simpson et al., 2000.) ж – жгутики, вб – вентральная бороздка, в которой проходит задний жгутик, к – кинетосомы, пк – передний микротрубоч- ковый корешок, лк – левый корешок с отходящими вторичными микротрубочками (вмт), пр – правый корешок с отходящими вторичными микротрубочками (вмт) и укрепляющей его фибриллой (ф), иф – исчерченная (комплексная) фибрилла, вв – внутренняя ветвь правого корешка, нв – наружная ветвь правого корешка с сопровождающими ее микротрубочками (мт), цф – цитофаринкс. Правая стенка вентральной бороздки на рис. Б отогнута наружу. Масштабная линейка на рис. А – 10 мкм.
112

лено. Общий список их (по: Patterson, 1999) приведен ниже с небольшими изменениями. Их изучение позволит решить многие таксономические проблемы и усовершенствовать систему протистов.
I. Свободноживущие гетеротрофные жгутиконосцы:
1. |
Acinetactis |
30. |
Metromonas |
2. |
Allantion |
31. |
Microcometes |
3. |
Allas |
32. |
Paramastix |
4. Alphamonas |
33. Paramonas |
||
5. Amphimonas |
34. Peltomonas |
||
6. |
Artodiscus |
35. |
Phanerobia |
7. Aulomonas |
36. Phloxamoeba |
||
8. Bodopsis |
37. Phyllomonas |
||
9. Bordnamonas |
38. Platytheca |
||
10. Campanoeca |
39. Pleurostomum |
||
11. Cladomonas |
40. Rhizomonas |
||
12. |
Clautriavia |
41. |
Proleptomonas |
13. |
Codonoeca |
42. |
Quadricilia |
14. Cyclomonas |
43. Rigidomastix |
||
15. |
Dallingeria |
44. |
Salpingorhiza |
16. Dimastigamoeba |
45. Schewiakoffia |
||
17. |
Dingensia |
46. |
Stenocodon |
18. Dinoasteromonas |
47. Stephanomonas |
||
19. Dinomonas |
48. Toshiba |
||
20. Diplocalium |
49. Trichonema |
||
21. |
Diplomita |
II.Паразитическиепротисты: |
|
22. |
Diploselmis |
||
23. Errera |
1. Amylophagus |
||
24. |
Fromentella |
2. |
Aphelidiopsis |
25. |
Heliobodo |
3. |
Barbetia |
26. |
Kamera |
4. |
Bertarellia |
27. |
Kiitoksia |
5. |
Bertramia |
28. |
Macappella |
6. |
Cibdelia |
29. |
Metopion |
7. |
Cingula |
113

ГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
8. |
Cristalloidophora |
45. |
Protomyxa |
9. Cytamoeba |
46. Pseudosporopsis |
||
10. Dinemula |
47. Rhabdospora |
||
11. |
Diplophysalis |
48. |
Rhinosporidium |
12. Ducelleria |
49. Rhyncodinium |
||
13. |
Echinococcidium |
50. |
Sergentella |
14. |
Ectobiella |
51. |
Serpentoplasma |
15. Elleipsisoma |
52. Spermatobium |
||
16. Embryocola |
53. Sphaerasuctans |
||
17. Endamoeba |
54. Spiriopsis |
||
18. Endemosarca |
55. Spirogregarina |
||
19. |
Endobiella |
56. |
Toxocystis |
20. Endomonas |
57. Trophosphaera |
||
21. Endospora |
58. X-клетки |
||
22. |
Eperythrocytozoon |
|
|
23. |
Globidiellum |
III. Водоросли: |
|
24. Gymnococcus |
1. Adinomonas |
||
25. |
Haematotractidium |
2. |
Àrchaeosphaerodiniopsis |
26. |
Hyalochlorella |
3. |
Aurospora |
27. |
Ichthyophonus |
4. |
Berghiella |
28. Immnoplasma |
5. Bjornbergiella |
||
29. Lymphocytozoon |
6. Boekelovia |
||
30. Lymphosporidium |
7. Camptoptyche |
||
31. Mononema |
8. Chalarodora |
||
32. Myrmicisporidium |
9. Chlamydomyxa |
||
33. Naupliicola |
10. Copromonas |
||
34. Neurosporidium |
11. Cyanomastix |
||
35. Ovicola |
12. Dinoasteromonas |
||
36. Palisporomonas |
13. Dinoceras |
||
37. |
Paradinemula |
14. |
Glaucocystopsis |
38. Paraplasma |
15. Goniodinium |
||
39. |
Parastasiella |
16. |
Heteromastix |
40. |
Physcosporidium |
17. |
Hillea |
41. |
Piridium |
18. |
Histiophysis |
42. |
Polysporella |
19. |
Isoselmis |
43. Protenterospora |
20. Melanodinium |
||
44. Protomonas |
21. Meringosphaera |
||
114

22. Monodus |
21. Elaeorhanis |
||
23. Nephrodinium |
22. Endalimax |
||
24. Pachydinium |
23. Enteromyxa |
||
25. |
Peliainia |
24. |
Flamella |
26. Petasaria |
25. Gymnophrydium |
||
27. Phialonema |
26. Hartmannina |
||
28. |
Pleuromastix |
27. |
Heterogromia |
29. |
Pseudoactiniscus |
28. |
Hyalodaktylethra |
30. Strobilomonas |
29. Iodamoeba |
||
31. |
Syncrypta |
30. |
Janickina |
32. |
Tetragonidium |
31. |
Kibisidytes |
33. |
Thaulirens |
32. |
Lagenidiopsids |
34. Thaumatodinium |
33. Leptophrys |
||
35. Thylakomonas |
34. Leukarachnion |
||
36. |
Triangulomonas |
35. |
Liegeosia |
|
|
36. |
Lithocolla |
IV. Амебоидные протисты: |
37. |
Malpighiella |
|
1. |
Actinocoma |
38. |
Martineziella |
2. Actinolophus |
39. Megamoebomyxa |
||
3. Aletium |
40. Myxodictyum |
||
4. Actinastrum |
41. Microgromia |
||
5. |
Actinelius |
42. |
Penardia |
6. Amphitrema |
43. Pleurophrys |
||
7. |
Apogromia |
44. |
Podactinelius |
8. Asterocaelum |
45. Podostoma |
||
9. Astrolophus |
46. Pontomyxa |
||
10. Balamuthia |
47. Protogenes |
||
11. |
Belaria |
48. |
Raphidiophryopsis |
12. |
Belonocystis |
49. |
Reticulamoeba |
13. Branchipocola |
50. Rhizoplasma |
||
14. |
Chamydophyrs |
51. |
Servetia |
15. Cichkovia |
52. Theratromyxa |
||
16. |
Cinetidomyxa |
53. |
Topsentella |
17. |
Clathrella |
54. |
Trizona |
18. Dictyomyxa |
55. Urbanella |
||
19. Dinamoeba |
56. Wagnerella |
||
20. |
Dobellina |
|
|
115

ГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
V. Протисты |
7. |
Ligniera |
|
неизвестной природы: |
8. Magosphaera |
||
1. |
Asthmatos |
9. |
Pansporella |
2. |
Endostelium |
10. |
Perkinsiella |
3. Euchitonia |
11. Phagomyxa |
||
4. |
Euglenocapsa |
12. |
Spongastericus |
5. |
Heliomonas |
13. |
Spongocyclia |
6. |
Hermisenella |
14. |
Spongospora |
116
ГЛАВА 3
Основные методы изучения строения клетки
Невооруженный глаз человека различает объекты в сотни микрон (рис. 3.1). Размеры большинства клеток протистов не превышают десятков микрон, и вместе с тем довольно сложно устроены. Поэтому для их изучения применяются различные приспособления, порой весьма сложные. В этой главе будет
кратко описана микроскопическая техника для их изучения и изложены основные методы.
Световая микроскопия
Световые микроскопы используются для наблюдений за клетками начиная с середины XVII века, т.е. уже около 350 лет. Они позволяют наблюдать объекты размером меньше бактерий, однако имеют предел разрешения, который определяется длиной световой волны. Степень разрешения микроскопа равна примерно половине длины волны используемого излучения. Длина волны видимого света око-
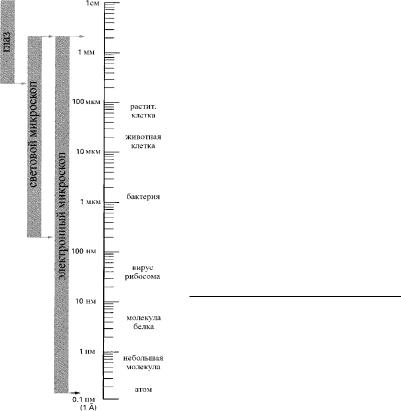
Рис. 3.1. Сравнение возможностей человеческого глаза и микроскопи- ческой техники с разной степенью разрешения.
Размеры клеток и их компонентов соотнесены с логарифмической шкалой.
1 ìì = 10-3 ì, 1 ìêì = 10-6 ì, 1 íì = 10-9 ì, 1Å (ангстрем) = 10-10 ì.
117

ГЛАВА 3. ОСНОВНЫЕ МЕТОДЫ ИЗУЧЕНИЯ КЛЕТКИ
ло 0,4 мкм, следовательно, степень разрешения любого светового микроскопа не может превышать 0,2 мкм. Другими словами, сколько бы мы ни увеличивали изображение объекта, мы не сможем различить 2 точки отдельными друг от друга, если расстояние между ними меньше 0,2 мкм (они будут восприниматься как одна точка). Для сравнения, толщина жгутика, или реснички эукариотной клетки, составляет 0,2 мкм, поэтому они плохо видны в световой микроскоп. Другая сложность заклю- чается в том, что клетки имеют небольшую толщину и обычно прозрачны для проходящего света. Получить более четкое изображение можно путем усиления контраста объекта. Это достигается двумя способами: специальное окрашивание изучаемых структур или применение дополнительных устройств (фазовоконтрастное, дифференционно-контрастное, электронные контрастирующие устройства).
Использование красителей
Основные красители были изобретены в конце позапрошлого века и с тех пор по настоящее время с успехом применяются в некоторых областях биологии и медицины. Это малахитовый зеленый, судановый черный, кумасси голубой, железный гематоксилин и некоторые другие. Их специфичность обычно невысока. Они позволяют выделять в клетке различным цветом углеводы, или нуклеиновые кислоты, или белки. Например, железный гематоксилин окрашивает ДНК, РНК и белковые структуры в черный цвет, реакция Фельгена выявляет только ДНК, окрашивая ее в ярко-красный цвет.
Для высокоспецифичных реакций, например для окраски макромолекул актина, в настоящее время используются специально выработанные против актина антитела, к которым присоединяются флюоресцентные метки, видимые в проходящем свете строго определенной длины волны. В процессе окрашивания меченые антитела избирательно соединяются с молекулами актина, и мы можем наблюдать распределение актиновых молекул во флюоресцентном микроскопе. Принцип работы
флюоресцентного микроскопа показан на рисунке 3.2. Флюоресцентные молекулы поглощают свет одной длины
волны, а генерируют свет большей длины волны. Поэтому, если
118
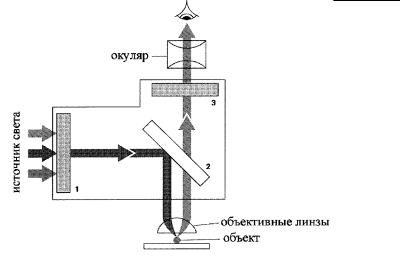
Рис. 3.2. Упрощенная схема строения современного флюоресцентного микроскопа.
Первый фильтр (1) пропускает свет одной длины волны (напр. 450–490 нм), который, отражаясь от дихронного зеркала (2), фокусируется при помощи объективной линзы на объекте. Здесь он возбуждает молекулы флюоресцентного красителя (напр. флюоресцина), который генерирует излучение большей длины волны (зеленый цвет, 520–560 нм). Свет от объекта проходит сквозь расщепляющее лучи зеркало (2), которое отражает свет с длиной волны меньше 510 нм и пропускает свет большей длины волны. Далее свет проходит через еще один фильтр (3), пропускающий излучение лишь с длиной волны 520–560 нм, т.е. зеленый свет, который мы и воспринимаем через окуляр.
поставить на пути отраженного от объекта света специальные фильтры, пропускающие свет только определенной длины волны, мы сможем увидеть свечение метки на темном поле. Обыч- но используются 2 вида флюоресцентных меток: флюоресцин, дающий ярко-зеленый свет при возбуждении (поглощении) голубым светом, и родамин, дающий интенсивный красный свет при возбуждении желто-зеленым светом.
В современных исследованиях часто используются так называемые двойные и тройные окраски, т.е. одна и та же клетка окрашивается двумя или тремя флюоресцентными метками раз-
119