

Протисты / Карпов С.А. Строение клеток протистов
.pdfГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
Рис. 2.47. Схема сложного жизненного цикла протостелиды Ceratiomyxella tahitiensis. (Ïî: Spiegel, 1990.)
1 – спорокарп, 2 – выходяшая из спорокарпа многоядерная амеба, 3 – амеба или участок пламодия с дегенерирующими ядрами, который превращается в зооцисту, 4–6 – три последовательных ядерных деления в зооцисте, 7–8 – формирование и выход из зооцисты 8 или менее жгутиковых клеток, 9–10 – превращение жгутиконосцев в амеб, 11 – плазмодий, 12 – фрагментация плазмодия на преспоровые клетки, 13 – растущий спороген.
Рис. 2.48. Схема жизненного цикла миксогастромицетов. (По: Sleigh, 1989.)
А – плодовое тело на стебельке, Б – выход из споры зооспор (В, Г) или миксамеб (Д), Е, Ж – стадия слияния миксамеб или зооспор с образованием зиготы (З), И – стадия молодого плазмодия, К – стадия сетчатого плазмодия (К), на основе которого формируются плодовые тела (Л).
90

Подкласс Dictyostelia Lister, 1909 Диктиостелиды (Ðèñ. 2.49)
Спорокарпы формируются на основе многоклеточного псевдоплазмодия.
Представители: Dictyostelium, Acytostelium, Polysphondilium.
Рис. 2.49. Жизненный цикл Dictyostelium discoideum. (Ïî: Carlile, Watkinson, 1994.)
1 – питающаяся амеба, 2
– стадия голодания с последующей агрегацией амеб, 3 – стадия псевдоплазмодия, 4 – миграция и дифференцировка псевдоплазмодия, 5 – формирование плодового тела, 6 – зрелая спора, 7 – выход амеб и их размножение, 8 – дифференцировка гамет и последующее слияние их клеток и ядер, 9 – стадия макроцисты, 10 – мейоз. n-гаплоидное ядро, 2n – диплоидное ядро.
Тип Haplosporidia Caullery & Mesnil, 1889 Гаплоспоридии (Ðèñ. 2.50)
Тканевые и целомические одноклеточные паразиты беспозвоночных животных. Обычно формируют одноклеточные и одноядерные споры, не имеющие полярной капсулы и полярного филамента. Трофическая стадия в тканях хозяина представляет собой одноили многоядерную клетку. Отличительная черта их цитоплазмы – наличие гаплоспоросом, которые участвуют в формировании оболочки в процессе превращения многоядерной клетки в споронт. Дальнейшие ядерные деления и увели- чение размеров тела сменяется множественным делением и образованием одноядерных споробластов. Пары споробластов
91
ГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
сливаются друг с другом, образуя двухъядерные клетки, за чем следует кариогамия. Клетки на этой стадии имеют форму песочных часов. Затем часть клетки с ядром (спороплазма) полностью окружается (заглатывается) безъядерной частью клетки (эписпоровая цитоплазма). Внутри эписпоровой цитоплазмы образуется чашевидная оболочка, отверстие которой закрыто крышечкой из плотного материала. Наружная поверхность оболочки имеет орнаментацию в виде оплетающих ее нитей.
Полностью жизненный цикл не изучен. Характерно нали- чие внутриядерного микротрубочкового веретена в интерфазных ядрах. Кристы в митохондриях трубчатые.
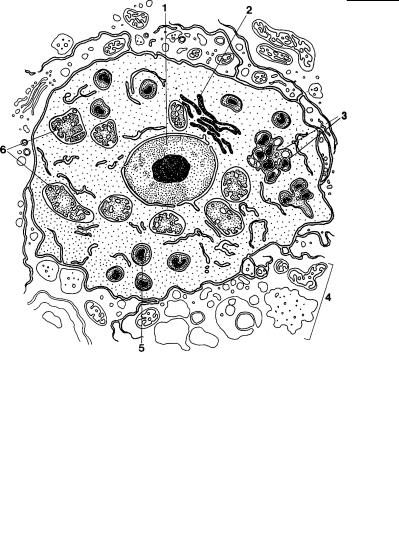
Рис. 2.50. Схема строения клетки гаплоспоридии. (По: Margulis et al., 1993.)
1 – ядро, 2 – аппарат Гольджи, 3 – зона формирования гаплоспоросом, 4 – органеллы клетки хозяина, 5 – гаплоспоросомы, 6 – митохондрии с пузырьковидными кристами.
92

Ранее гаплоспоридий считали классом в типе Ascetospora. Первые данные молекулярной филогении по генам 18S рРНК указывали на их близость к альвеолатам и даже непосредственно инфузориям, но последние комбинированные данные по сиквенсам генов актина и рРНК малой субъединицы показывают, что эта группа формирует независимый кластер.
Всего известно 3 рода: Haplosporidium (23 âèäà), Minchinia (4 âèäà), Urosporidium (6 видов).
Тип Foraminifera D’Orbigni, 1826 Фораминиферы (Ðèñ. 2.51)
Трофонт окружен однокамерной или многокамерной раковинкой, построенной на органической основе. Камеры раковинки сообщаются между собой через специальные отверстия – форамены. Раковины имеют одно, два или несколько отверстий – устьев, служащих местом выхода нитевидных псевдоподий – гранулоретикулоподий, образующих вокруг раковины ловчую сеть (ретикулум). Жизненные циклы с чередованием полового и бесполого поколений, с промежуточной редукцией. Гаметы с 1–3 жгутиками или вторично амебоидные. Обычно многоядерные, часто с гетероморфными ядрами. Преимущественно морские. Есть свободноживущие и паразитические виды. В типе на- считывается около 40 000 видов. Подавляющее большинство (около 80%) представлено ископаемыми формами, которые найдены в отложениях Кембрий – Голоцен. Филогенетические связи неясны. Последние данные молекулярной филогении указывают на их близость к альвеолатам.
Представители: Lingulina, Globigerina, Rhabdammina.
Тип Heliozoa Haeckel, 1866 Солнечники (Ðèñ. 2.52)
Гетеротрофные обычно радиально симметричные протисты с длинными радиальными аксоподиями, которые выходят из одного центра организации (аксопласта или центропласта). Микротрубочки в аксоподиях расположены в строгом порядке. В клетке нет центральной капсулы, внутренний скелет часто отсутствует или образован органическими или кремниевы-
93

ГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
94
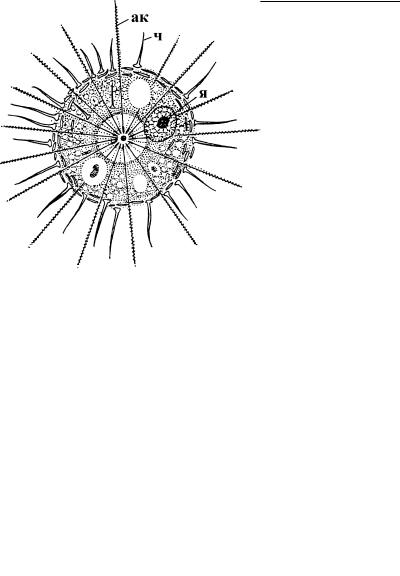
 Рис. 2.51. Схема жизненного цикла фораминиферы Cibicides lobatulus. (По: Михалевич, 1999.)
Рис. 2.51. Схема жизненного цикла фораминиферы Cibicides lobatulus. (По: Михалевич, 1999.)
1 – гаметы; 2 – сливающиеся гаметы образуют зиготу; 3,4 – однокамерные 2- и 3-ядерные агамонты; 5-8 – растущий многокамерный агамонт с макронуклеусом (ма) и микронуклеусами (ми), мигрирующими от центральной камеры к периферии; 9 – вышедшие из цисты агаметы; 10-13 – растущие гамонты. R1 и R2 – редукционные деления, га – гаметы внутри раковинки гамонта, ц – циста вокруг агамонта с дегенерировавшим макронуклеусом, я – ядра.
ми иглами. На поверхности клетки многих видов имеются че- шуйки различного размера и формы. Фибриллярные скелетные образования не встречаются. Экструсомы представлены мукоцистами и кинетоцистами. Кристы в митохондриях, за редкими исключениями, пластинчатые. Имеются трофонты со жгутиками. Преимущественно пресноводные. После выведения из этой группы актинофриидных солнечников она стала более однородной, но полифилия не исключается.
Рис. 2.52. Внешний вид центрохелидного солнечника. (По: Кусакин, Дроздов, 1998.)
ак–аксоподии, ч–чешуйки на поверхности клетки, я–ядро.
95

ГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
Класс Axoplasthelidea Febvre-Chevalier, Febvre, 1983. Аксопластидные солнечники
Центр организации микротрубочек аксоподий представлен аксопластом.
Представители: Actinocorine, Gymnosphaera, Hedraiophrys.
Класс Centroplasthelidea Febvre-Chevalier, Febvre, 1983. Центропластидные солнечники
Центр организации микротрубочек аксоподий представлен центропластом.
Представители: Heterophrys, Acanthocystis, Raphidiophrys.
Радиолярии
Класс Acantharia Mьller, 1855 Акантарии (Ðèñ. 2.53)
Радиально-симметричные протисты с аксоподиями и внутренним минеральным скелетом из игл, образованных солями стронция. В клетке хорошо развит эктоплазматический кортекс и сократимые фибриллы - мионемы. Аксонемы сходятся к центральному аксопласту; в центре клетки соединяются и скелетные иглы. Типичные формы имеют 10 диаметральных или 20 радиальных игл со стабильным положением. Имеются цисты и
Рис. 2.53. Внешний вид акантарии
Xiphacantha alata. (По: Шевяков, 1926.) ак–аксоподии, èã–èãëû, ми– мионемы.

многоядерные формы. В ископаемом состоянии не сохраняются. По очень немногим данным молекулярной филогении, группа весьма обособлена.
Представители: Holacantha, Amphilonche, Acanthometra.
Класс Polycystina Ehrenberg, 1838 Полицистины (Ðèñ. 2.54)
Протисты с радиальными аксоподиями и ажурным наружным кремниевым скелетом, который хорошо сохраняется в ископаемом состоянии. Центральная капсула образована гликопротеиновыми пластинками и снабжена специализированными зонами – фузулами, через которые микротрубочки аксоподий выходят наружу. Центры организации микротрубочек аксоподий (аксопласты) могут располагаться снаружи центральной капсулы (у фузул) или внутри нее. Цитоплазма снаружи от центральной капсулы сильно вакуолизирована и часто заселена симбиотическими водорослями. Митохондрии с трубчатыми кристами. Расселительные двухжгутиковые стадии содержат кристаллы сульфата стронция. По очень немногим данным молекулярной филогении, являются сестринской группой энтамеб.
Представители: Thalassicolla, Arachnosphaera, Plagiocarpa.
Рис. 2.54. Внешний вид скелета полицистины
Hexacontium asteracanthion. (Ïî: Sleigh, 1989).
97

ГЛАВА 2. КРАТКАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ ТАКСОНОВ ПРОТИСТОВ
Класс Phaeodaria Haeckel, 1879 Феодарии (Ðèñ. 2.55)
Группа морских радиально-симметричных протистов с аксоподиями, имеющих полый скелет из кремнезема. Центральная капсула в виде билатерально-симметричного мешочка. В стенке центральной капсулы есть 3 специализированные зоны: апикальное астропиле (цитофаринкс) и два парапиле, в которых находятся аксопласты аксоподий. Характерно наличие желто-коричневой аморфной массы – феодия, расположенного рядом с астропиле. Ископаемых форм почти нет.
Представители: Phaeodina, Aulacantha, Castanarium.
Рис. 2.55. Внешний вид феодарии
Aulospathis variabilis triodon.
(По: Решетняк, 1966.) иг – скелетные иглы, ф – феодий, цк – центральная капсула.
Ризоподы
Класс Lobozea Carpenter, 1861 Лобозные амебы (Ðèñ. 2.56)
Амебоидные протисты с лобозными псевдоподиями. Если формируют филозные псевдоподии, то они образуются из более широкой гиалиновой лобоподии. Особи обычно одноядерные, редко двухили многоядерные, иногда встречается истин-
98

Рис. 2.56. Внешний вид некоторых лобозных амеб. (По: Margulis et al., 1993.)
А – полиподиальная голая амеба, Б – моноподиальная амеба, В – раковинная амеба.
ный плазмодий. Плодовых тел не образуют. Митохондриальные кристы трубчатые. Тело клетки без раковинки (отряд Gymnamoebia Haeckel, 1862) или имеет раковинку (отряд Testacealobosia Saedeleer, 1934). Свободноживущие, редко паразитические. Населяют морские и пресные водоемы, почву. Очень немногие молекулярные данные указывают на полифилетичность группы.
Представители: Amoeba, Mayorella, Arcella.
99