

Сафонов А.И. Эволюционное учение. Классики и современники
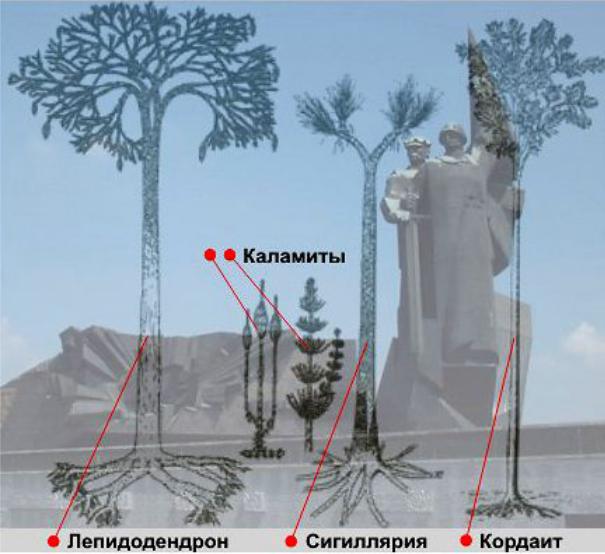
.pdfИтак, мы видим роскошные 20-40 метровой высоты стволы плауновидных (лепидодендронов, ботродендронов и сигиллярий) – рис. 116, кордаитов с их метровыми лентообразными листьями на вершинах, чуть пониже – членистостебельные (каламиты) и множество папоротников от травянистых до древовидных в 6 -7 метров высоты.
Еще можно увидеть грибы у подножия деревьев, напоминающие наши шампиньоны, мхи, лишайники да почву, устланную одеялом из листьев кордаитов и коры лепидодендронов, наподобие подстилки из хвойных иголок в современных лесах.
Вообразите на минуту, что на месте, где сегодня раскинулся миллионный Донецк, высятся тропические двадцати, а то и сорокаметровые стволы плауновидных, хвощеобразных и папоротниковых, залитых водой, а между ними носятся стрекозы с размахом крыльев метровой ширины.
Рис. 116 – Пейзаж каменноугольного периода
А вот описание карбоновых болот из замечательной книги К. Еськова, российского палеонтолога: «Теплый и влажный климат господствовал на материках длительное время. Все это создало чрезвычайно благоприятные условия для развития наземной флоры, в том числе высших растений каменноугольного периода – кустов, деревьев и травянистых растений, жизнь которых была тесно связана с водой. Они росли главным образом среди громадных болот и озер, близ солоновато водных лагун, на побережье морей, на влажной илистой почве и чем-то походили на современных мангровых, которые растут на низинных берегах тропических морей, в устьях больших рек, в болотистых лагунах, поднимаясь над водой на высоких корнях-ходулях.
В самом начале карбона на континентах начинается угленакопление – а это свидетельствует о том, что на пути стока вод стояли мощные растительные фильтры. Не будь их, остатки растений непрерывно смешивались бы с песком и глиной, так что получались бы обломочные породы, обогащенные растительными остатками – углистые сланцы и углистые песчаники, а не настоящие угли». На что были похожи каменноугольные ландшафты – предшественники нынешних донецких степей? Вспомним, что на протяжении многих миллионов лет Донецкий бассейн представлял собой глубокий залив, периодически осушавшийся и вновь затоплявшийся морскими водами. Кстати, водами Уральского океана, омывавшего в ту эпоху восточные берега континента Балтика. Заболоченные низины сменяли невысокие поднятия, устья рек, переходящие в морское мелководье – своего рода леса-водоемы (лепидофиты).
Продолжаем описание Еськова: «Лепидофитовые «леса-водоемы» занимали выположенные приморские низменности и именно в этих приморских бассейнах шли основные процессы углеобразования. Специалисты по лесоведению предполагают, что у лепидофитов естественное выпадение подроста происходило (в отличие от современных
441
лесов) не на ранних стадиях его роста, а на поздних: то есть сперва вырастает густая поросль, а потом она одновременно падает. Эти неритмичные поступления больших количеств мертвой органики в детритную цепь экосистемы («то густо, то пусто») создавали существенные проблемы с утилизацией этой органики. С другой стороны, противоэрозионные свойства этих «лесов», по сравнению с современными, были весьма слабы, а водоразделы продолжали размываться почти столь же интенсивно, что и прежде. Доводом в пользу отсутствия (или очень слабого развития) растительности на водоразделах являются очень частые в это время прижизненные захоронения деревьев вместе с их корневыми системами: такое случается при быстром выносе большой массы осадков (вроде селевого потока), которая «заживо погребает» растения. В итоге большой отпад в карбоновых «лесах-водоемах» (рис. 117) при высокой эрозии (обеспечивающей его быстрое захоронение) приводил к тому, что значительная доля неокисленного органического углерода безвозвратно уходила из экосистемы и превращалась в запасы каменного угля.
Рис. 117 – Символизм эпох г. Донецка
Появлению обширных залежей каменных углей способствовали жаркий и влажный климат, обилие древовидных растений, произрастающих на заболоченных почвах, а также особенности произрастания лепидофитовых лесов («леса-водоемы»).
Что до лепидофитовых лесов каменноугольного периода, то их экологическая структура была крайне своеобразна и не имеет современных аналогов. Это были неглубокие (первые метры), переполненные органическими остатками водоемы, а формировавшие эти «леса» высокоствольные (до 40 метров) плауновидные и хвощеобразные были гелофитами,
442

стоявшими «по колено в воде». С другой стороны, это были и не болота современного типа, где деревья вырастают поверх торфяника; в карбоновом «лесу» корневые системы лепидофитов (стигмарии) располагались ниже торфоподобной органической массы, а сами деревья прорастали сквозь нее и многометровый слой «валежника» (см. рис. 118).
а
б
Рис. 118 – Окаменелости: а) характерный пример лесной подстилки – смеси коры листьев, мелких веточек, споровых шишек и пр. мелоч; отвал шахты им. Шверника;
б)отпечаток листа кордаита, отвал шахты Заперевальная
Основные фотосинтезирующие поверхности этих древовидных споровых составляли не листья (довольно жалкая «метелка» на самом кончике 40-метровой «палки»), а периодически опадающая толстая зеленая кора; по аналогии с современными листопадными лесами, эти карбоновые леса иногда называют «коропадными».
443
В этой подстилке из опавшей коры существовала богатая фауна членистоногих
– паукообразных и многоножек; среди последних попадались гиганты длиною до
метра. |
Именно |
растительноядные |
|
двупарноногие |
многоножки |
были |
|
основными (а может – и единственными) в то время потребителями опавшей коры. Описываемые растительные сообщества росли на каменноугольных болотах, располагавшихся в низинах, вдоль рек и береговой линии моря. Что касается возвышенных, умеренно засушливых участков ландшафта, то там господствовали первые хвойные (валчиевые) и кордаитовые. Наиболее совершенные растения карбона – кордаиты. Их цилиндрические безлистые стволы (до 40 м) с высотой разветвлялись. Ветви имели на концах широкие линейные или ланцетные листья с сетчатым жилкованием. Мужские спорангии имели вид почек. Из женских спорангиев развивались ореховидные плоды.
Результаты микроскопического исследования плодов показывают, что эти растения, похожие на цикадовых, являлись переходными формами к хвойным растениям.
Фауна каменноугольного периода. Что ползало, летало и плавало?
(рис. 119-126)
Слово Кириллу Еськову: "На суше это, безусловно, пауки и скорпионы, перешедшие в каменноугольный период из более ранних эпох. Длина скорпионов достигала 25 сантиметров, а огромные многоножки (артроплевриды) вымахали в длину на 2 м и вышагивали на трехстах парах ног. Как уже отмечалось ранее, в каменноугольном периоде жизнь насекомых протекала в похожей на валежник подстилке из опада крупных листьев и коры деревьев.
Там резвились тараканы по 10 см длиной, разнообразные многоножки и паукообразные".
Описываемая «крупноразмерность» насекомых в карбоне требует отдельного пояснения.
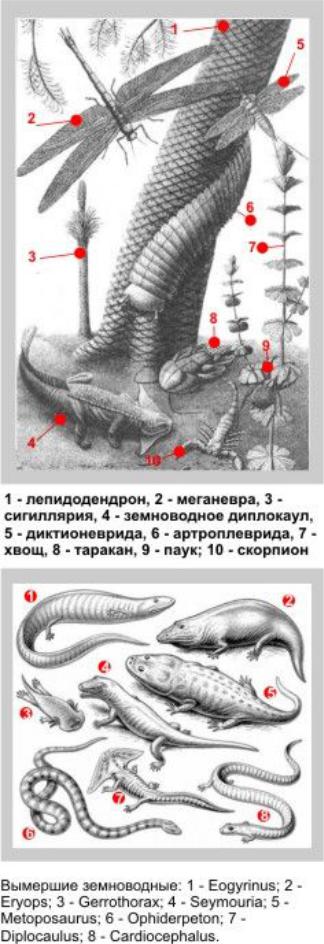
Рис. 119 – Растения и животные каменноугольного периода
444

Все дело в том, что концентрация кислорода в атмосфере того периода возросла до 35% в отличие от современных 21%. И если в конце девона концентрация кислорода составляла около 15%, то ее повышение до столь высокого уровня должно иметь на то вескую причину. (Если бы содержание кислорода перевалило за 35%, вся наземная органика самовозгорелась бы. Впрочем, даже при таком его уровне с середины каменноугольного периода участились лесные пожары).
Произошло это вследствие появления в девоне первых деревьев. Их широкое распространение по планете в карбоне способствовало усилению процессов фотосинтеза, в результате которого потреблялся углекислый газ атмосферы (связывался углерод), а вода расщеплялась на водород и кислород. В принципе, процесс фотосинтеза балансирует соотношение кислород – углекислый газ. Углерод усваивается деревьями, а кислород попадает в атмосферу и участвует в окислении отмершей древесины.
Однако, как мы помним, в карбоне значительная часть растительной массы консервировалась в прибрежных зонах, чтобы со временем превратиться в уголь, т.е. достаточно много кислорода оставалось не использованным и, соответственно, нарушался баланс. По одной из гипотез американских ученых причина повышения концентрации кислорода заключается в том, что деревья каменноугольного периода имели более толстую кору, которая содержит много лигнина, т.е. отмиравшие стволы состояли в большей степени из лигнина, чем из целлюлозы. Лигнин же, как известно, разлагается хуже, чем целлюлоза и, таким образом, тратится значительно меньше кислорода для окисления органики.
Теперь о самих насекомых. «Кровь» насекомых (гемолимфа) лишена дыхательных пигментов и не участвует в переносе кислорода. Дыхание же осуществляется при помощи трахей – ветвящихся трубочек, непосредственно соединяющих клетки внутренних органов с воздушной средой. Приток кислорода внутрь тела (так же, как отток углекислого газа) происходит за счет диффузии при разнице парциальных давлений этих газов на внутреннем и внешнем концах трубки. Такой механизм подачи кислорода жестко ограничивает длину трахейной трубки, поэтому максимальный размер тела самого насекомого не может превышать двойной длины трахейной трубки. Поэтому существование в карбоне таких гигантов, как стрекоза-меганевра или основная ее добыча – диктионевриды размером с голубя, объясняется тем, что в те времена парциальное давление кислорода в земной атмосфере было заметно выше, чем ныне – как оно и должно быть при захоронении больших количеств неокисленного углерода. Кстати, видимо именно это увеличение соотношения О2/СО2 в атмосфере планеты вызвало «обратный парниковый эффект», приведший к Гондванскому оледенению. К концу каменноугольного периода среди насекомых появились хищники с внешними хватательными органами (стрекозы и прямокрылые). Самыми крупными из них были стрекозы (рис. 120) с подвижной головой, несущей огромные глаза и режущие челюсти» (меганевра).
Рис. 120 – В левой части иллюстрации – отпечаток меганевры из музея Луганского университета, в правой – реконструкция насекомого с сайта ВВС.
445
Не стану утруждать себя детальным описанием насекомых карбона, это до меня прекрасно сделали ряд авторов-палеонтологов, работы которых здесь время от времени цитируются. Например А.Ю. Журавлев в своей публикации пишет: «Благодаря повышенной концентрации кислорода, стрекозы размахнули крылья на 70 см, диктионевриды – более, чем на 40 см. Диктионевриды (греч. «сетчатожилкованные») были каменноугольными насекомыми с колющим хоботком, крупными расправленными крыльями и парой длинных членистых хвостовидных придатков (чем напоминают современных поденок). Прекрасные летуны с клювовидной головой они протыкали и высасывали жестким колющим хоботком семязачатки кордаитов и других растений. Необходимо заметить, что появление в конце раннего карбона крылатых насекомых вообще было одним из ключевых событий в истории жизни на суше. Собственно говоря, вся история наземных экосистем – это фактически история трех групп живых организмов: сосудистых растений, насекомых и позвоночныхтетрапод, а также их взаимодействия между собой. Оговорка насчет «крылатых» насекомых не случайна: вообще-то первые насекомые появились в девоне, однако именно с возникновением способности к полету у крылатых насекомых эта группа стала самой процветающей в животном царстве.
Большая плотность воздуха обеспечила достаточную подъемную силу и условия для полета. И не только для планирования. Машущий полет требует постоянного и скорого обновления кислорода в организме, что возможно при высоком атмосферном давлении и быстром обновлении газа в трахеях. Крылья развились у крупных прыгающих насекомых из боковых пластинчатых выростов, служивших для контроля положения в воздухе. У каменноугольных диктионеврид было даже три пары пронизанных жилками и покрытых щетинками крыловидных выростов. Но двигались только две последние пары. С такими крыльями трудно было спрятаться, но можно было улететь помимо своего желания с порывом ветра. С лесом связано становление наиболее многочисленной группы животных насекомыхшестиножек. Небольшие размеры позволяли им проживать в очень ограниченном пространстве (почве, растениях и их остатках, на животных). Насекомые смогли превратить в пищу практически любое живое и мертвое органическое вещество, включая мало-мальски перевариваемую и совершенно непитательную древесину. Первые насекомые довольствовались грубым измельчением спор и семязачатков челюстями. Вопреки всем ухищрениям многоножек, клещей и насекомых зеленая масса растений оставалась недоупотребленной и уходила в опад, а оттуда практически прямо в уголь».
Признаки «четвероногости» и «земноводности» независимо накапливались в разных ветвях кистепёрых рыб в течение всего девонского периода.
Это были легочное и кожное дыхание, трехкамерное сердце и смешанное кровообращение, способность выделять мочевую кислоту вместо аммиака (для выведения лишнего азота, что обеспечивало более разумный расход воды) и опорная конечность. У двоякодышащих рыб тоже появилось трехкамерное сердце из плавательного пузыря.
Но морские кистепёрые рыбы обрели больше земноводных признаков, чем пресноводные двоякодышащие и опередили их, заняв это многообещающее для продвижения по эволюционной лестнице место.
Первые, еще девонские, тетраподы – примитивные амфибии лабиринтодонты (название это происходит от их зубов с лабиринтоподобными складками эмали – структура, прямо унаследованная от кистеперых), такие как ихтиостега и акантостега, в захоронениях всегда встречаются вместе с рыбами, которыми, судя по всему, и питались. Например, акантостега, обитала в болотах, располагавшихся на территории современной Восточной Гренландии, примерно 360 млн. лет назад. Благодаря четырем конечностям животное поднимало голову из воды и дышало атмосферным воздухом, но выбраться на берег оно еще не могло. Лабиринтодонты (рис. 121) были покрыты чешуей, как рыбы, имели хвостовой плавник (похожий на тот, что мы видим у сома или налима), органы боковой линии и – в некоторых случаях – развитый жаберный аппарат; конечность их еще не пятипалая (число пальцев достигает 8), и по типу сочленения с осевым скелетом – типично плавательная, а не опорная.
446

Все это не оставляет сомнения в том, что существа эти были чисто водными; если они и появлялись на суше при неких «пожарных» обстоятельствах (пересыхание водоема), то компонентом наземных экосистем наверняка не являлись.
Рис. 121 – Реконструкция древних животных Донбасса
Ситуация принципиально изменилась с начала карбона, когда в уже знакомых нам «лесах-водоемах» возникла богатая фауна из амфибиотических и наземных членистоногих – паукообразные, многоножки, а потом и насекомые. Среди карбоновых амфибий появляются небольшие (менее 1 м), похожие на современных саламандр антракозавры, имеющие несомненные приспособления к наземной жизни и явно ориентированные на питание беспозвоночными (а не рыбой – как их более крупные родственники). Именно антракозавров считают предками рептилий. Первые рептилии – карбоновые капториниды – напоминали по
447

внешнему виду крупных ящериц, причем, судя по строению их челюстного аппарата, они специализировались именно на питании насекомыми.
Название «антракозавр» означает «угольная ящерица», поскольку их остатки часто встречаются в угольных пластах. Конечности у этих короткотелых животных были развиты, но недостаточно для передвижения по суше. Они так и остались водными обитателями, даже
схвостовым плавником, и вымерли к середине триасового периода.
Впозднекаменноугольную эпоху существовали полуметровые тупорылые потомки антракозавров сеймурии. Будучи личинками, они сохраняли жаберные дуги, но взрослые выглядели настоящими пресмыкающимися: голова поворачивалась относительно туловища, легкие всасывали воздух, а органы слуха слышали звуки, разносимые в воздушной среде. Отпала необходимость в кожном дыхании, и чешуя покрыла тело, защищая от высыхания, гибельного для земноводных.
Вкаменноугольном периоде появляются крокодиловидные лабиринтодонты. Это были похожие на лягушек (своих возможных потомков) и одновременно на крокодилов земноводные с большими глазами и длинным рылом. Их личинки с короткой головой, длинным хвостом, хрящевым скелетом и внешними жабрами напоминали головастиков с лапками. Среди лабиринтодонотов встречались настоящие гиганты до полутора метров длиной, с головой в треть тела. Из нёба и нижней челюсти у них торчали клыкообразные зубы. Такие живые капканы скрывались на дне, прикидываясь бревном, и выжидали, кто заплывет в широкий лягушачий рот.
Земноводные отличались большим разнообразием формы и размеров. Большую группу составляли амфибии до метра длиной, хотя преобладали в основном мелкие (около 30 см). Часто они теряли лапки, превращаясь в змеевидных существ, и обладали различной величиной и формой тела: одни походили на тритонов или ящериц, другие на крокодилов и змей. Их тело было покрыто панцирем, состоявшим из мелких чешуек или палочек. У некоторых форм значительных размеров панцирь состоял из больших толстых костных щитов. Голова была плоской, большею частью треугольной, пасть была вооружена острыми зубами.
Позднекаменноугольный диплокаул – рис. 122 (греч. «двойная рукоятка») щеголял треугольным черепом с далеко выступавшими углами (глаза и ноздри сверху, маленький рот снизу).
Рис. 122 – Диплокаул
448
К концу каменноугольного периода земноводные стали крупнейшими хищниками, несколько потеснив скорпионов, пауков, многоножек и насекомых. Впрочем, из-за пассивного дыхания тело на воздухе скоро остывало и теряло влагу, что привязывало земноводных к водной среде. Совмещение органов глоточного дыхания и питания мешало развитию челюстей и легких. Рот открывался широко, но «неудачно» расположенные челюстные мускулы не создавали значительного давления. Голова казалась вросшей в туловище. Земноводные приспособились к пассивной охоте по системе «живой капкан», но не могли преследовать свои жертвы. Не приспособились они к растительной пище. Господство их было недолгим.
Чем объяснить итоговый эволюционный неуспех амфибий? Ведь этот класс процветал только в карбоновом периоде (причем – только на территории тогдашних тропиков, в Еврамерике), когда у него не было конкурентов; во все же остальные времена они занимали сугубо подчиненное положение в экосистемах – как наземных, так и пресноводных. Судя по всему, амфибий сгубила даже не обязательная для них связь с водой (они не выработали яйца и размножались икрой), а неисправимое несовершенство их дыхательной системы. Слабость легочного дыхания не позволяет амфибиям освободить от дыхательных функций кожу. Об этом обычно упоминают в той связи, что земноводные должны всегда сохранять покровы влажными (именно в этой влаге и растворяется воздушный кислород, «всасываемый» затем кожей), а потому они навсегда прикованы к воде. Не освободив от дыхательных функций кожу, невозможно изолировать друг от друга большой и малый круги кровообращения и отделить венозную кровь от артериальной. У амфибий сердце трехкамерное – два предсердия и желудочек – и кислород приносят в сердце вены, идущие не только от легких (малый круг) но и от кожи (большой круг), а кровь в сердце всегда будет смешанной – артериально-венозной. Пока же кровь не разделена на артериальную и венозную, уровень энергетического обмена организма будет принципиально невысоким. К тому же при постоянно влажных покровах любое повышение температуры тела будет тут же сводиться на нет за счет поверхностного испарения – термическое охлаждение. Все это полностью закрывает амфибиям путь к достижению любых вариантов эндотермии (теплокровности) – с понятными эволюционными последствиями.
Из-за единственно доступного для амфибий способа наполнения легких («проглатывания» воздуха) необходимо сохранять «подчелюстной насос», поднимающий и опускающий дно ротовой полости. Размещение последнего требует совершенно специфической «лягушачьей» формы черепа – очень широкой и приплюснутой. При этом челюстная мускулатура оказывается размещенной крайне невыгодным образом: ее можно подсоединить лишь к самому челюстному суставу с задней его стороны. Для того, чтобы развивать при помощи таких челюстей сколь-нибудь приличные усилия, приходится наращивать объем челюстной мускулатуры; в итоге эта мускулатура занимает практически все пространство черепа, попросту не оставляя места для мозговой коробки. То есть амфибии – существа принципиально «безмозглые», и это закрывает им путь к сложным формам поведения.
Земноводных превзошли пресмыкающиеся. История пресмыкающихся началась в среднекаменноугольную эпоху. В то время кора у плаунов была толстой и прочной, а сердцевина сгнивала. В дуплистых стволах прятались различные животные. К середине каменноугольного периода возникли мелкие (около 10 см длиной) ящерицеподобные капториноморфы. Древнейшие капториноморфы найдены все в тех же плауновых дуплах. Там они выискивали насекомых и улиток, которых давили многочисленными притупленными цилиндрическими зубами. От этих вполне наземных существ с длинными конечностями и развитой шейной мускулатурой произошли прочие пресмыкающиеся пеликозавры (греч. «парусные ящерицы») и зверообразные (предки млекопитающих).
Для потребления растительной пищи необходимы крепкие режущие и секущие зубы, размельчающие ее перед глотанием, вместительный желудок и помощь симбионтов в переваривании. На такую пищу смогли перейти крупные пресмыкающиеся. В силу своих
449

размеров они обладали достаточно мощными челюстями и повышенной температурой внутри тела. Покрытое известковой скорлупой яйцо появилось у пресмыкающихся задолго до курицы и надежно решило проблему сохранения влаги.
К концу каменноугольного периода в низинных участках экваториального лесах появились крупные пресмыкающиеся. Первыми были грузные (3-4 м длиной) пеликозавры эдафозавры (греч. «почвенная ящерица»). Эдафозавры, ползавшие на коротких конечностях, могли питаться низкорослой наземной растительностью, ракушками и насекомыми. У всех пеликозавров был парус – кожная перепонка, поддерживаемая отростками спинных позвонков и пронизанная кровеносными сосудами.
Рис. 123 – Из коллекции Олега Крисака, ДонНТУ (найдено в Донбассе)
Рис. 123 – а) найдено в карьере п. Ларино под Донецком, б) найдено на отвале шахты Куйбышевская, Донецк
Морской животный мир карбона характеризовался разнообразием видов. Одиночные кораллы нередко имели толстые известковые стенки, колониальные кораллы образовывали рифы. В это время интенсивно развиваются иглокожие, в частности морские лилии и морские ежи. Многочисленные колонии мшанок иногда образовывали мощные известняковые отложения. Чрезвычайно развились плеченогие моллюски, в частности продуктусы. Величина их раковин достигала 30 см в поперечнике. Одна створка раковины
450