

Сафонов А.И. Эволюционное учение. Классики и современники
.pdfГлава 6 Новые виды, или
Как предотвратить скрещивание
Вид – одно из самых странных и спорных понятий в биологии. Оно было введено английским ботаником Джоном Рэем в XVII веке. Вид, определенный Рэем, представлял собой череду неизменных предков и потомков, причем родитель мог дать начало только такому же, как и он сам, организму. Рэй наблюдал на своем огороде, что из семян культурного растения может вырасти только то же самое растение, а сорняки вырастают из других, своих собственных семян или корневищ (в то время многие думали, что они появляются из тех же семян культурных растений нежелательным, хотя и неизбежным приращением). Так что исходное определение вида такое: совокупность тождественных друг другу организмов, способных давать подобное себе потомство.
Через несколько промежуточных ступеней концепция вида обрела помимо свойства устойчивости еще и свойство изменчивости. С одной стороны, очевидно, что в природе действительно существуют более или менее четко обособленные совокупности похожих особей, которые, как правило, могут скрещиваться и производить плодовитое потомство и которые связаны более тесным родством друг с другом, чем с представителями других подобных совокупностей. С другой стороны, особи в группе никогда не бывают на одно лицо, как двое из ларца. С третьей, далеко не всегда удается четко и однозначно определить, где проходит граница, какие группы особей следует считать самостоятельнымии видами, а какие – только подвидами, породами или разновидностями. Поэтому попытки дать виду строгое определение, избавиться от всех «более или менее» и «как правило» неизменно терпят неудачу. Предложены десятки «концепций вида», сотни критериев, тысячи определений – все без толку. Понятие как было расплывчатым и нечетким, так и осталось.
Удивляться тут нечему. Как отмечал еще Дарвин, виды не должны, да и не могут быть совершенно четкими и дискретными. Ведь их главное свойство – пластичность, способность постепенно меняться в силу законов изменчивости, наследственности и отбора. Между разновидностью и видом нет четкой грани, потому что разновидности со временем могут стать видами, а существующие сегодня виды в прошлом были всего лишь разновидностями каких-то других, предковых видов. В современной биосфере мы застали разные совокупности особей на разных стадиях этого процесса. По-видимому, периоды относительно стабильного существования вида чередуются с периодами быстрых изменений, причем первые продолжаются в среднем дольше. Именно поэтому в биосфере все-таки преобладают «хорошие», четко обособленные виды, в то время как спорные, переходные случаи встречаются реже. Хотя и это тоже спорно. Трудно найти такую группу животных или растений, в классификации которой не было бы сомнительных мест.
При всей расплывчатости формального определения виды все же реальны, по крайней мере у организмов, размножающихся половым путем. Вид – это прежде всего единый генофонд, совокупность аллелей, эволюционирующих как одно целое (см. главу 3). С бесполыми организмами сложнее. Непонятно, как провести грань между разновидностью и видом, например, у бделлоидных коловраток. Как бы мы ни старались, и то и другое – всего лишь совокупности более или менее родственных, конкурирующих друг с другом клонов, неспособных к регулярному обмену наследственным материалом. Тем не менее специалисты по коловраткам изо всех сил стараются (честно говоря, не слишком успешно) поделить на четкие виды как моногононтных коловраток, размножающихся половым путем, так и бесполых бделлоидных – и поди разбери, что при этом имеется в виду на самом деле.
–––––
Эволюция видов и языков: единство постепенности и дискретности
Эволюция языков похожа на эволюцию биологических видов, хотя это сходство не следует преувеличивать. Лингвистическая эволюция в основном нейтральна – не имеет приспособительного характера. Нельзя утверждать, например, что грамматика чукотского языка лучше приспособлена к холодному
241
климату, чем грамматика языков африканцев. В биологической эволюции, впрочем, тоже велик элемент нейтральности. Для построения эволюционных деревьев биологи предпочитают использовать именно нейтральные признаки. Дело
втом, что приспособительные признаки – менее надежное свидетельство родства. Они могут возникать у неродственных форм в сходных условиях из-за одинаковой направленности отбора. Специалисты по исторической лингвистике активно и успешно используют математические методы построения эволюционных деревьев,
разработанные биологами (Lieberman et al.; Pagel et al., 2007).
Аналогия с эволюцией языков помогает лучше понять два ключевых свойства биологической эволюции (на первый взгляд противоречивых): постепенность изменений и относительную дискретность видов.
Спостепенностью все просто. Языки, как и виды, эволюционируют путем накопления мелких изменений. Один язык (например, латынь) превращается в другой (например, итальянский) постепенно. Каждое следующее поколение говорит лишь немного иначе, чем предыдущее. Не бывает так, чтобы родители, говорящие на латыни, родили детей, которые, научившись говорить, вдруг заговорили по-итальянски. Так же и в эволюции видов: родители одного вида не могут родить детенышей, относящихся уже к другому виду (в одном научно-популярном фильме были кадры, в которых мамочка из породы питекантропов родила сапиентного детеныша и с удивлением его разглядывает. Такого, конечно, не бывает). Переход происходит плавно и незаметно. Четкие различия проявляются, только если сравнивать конечные звенья длинной цепи постепенных изменений.
Менее очевидны причины дискретности. Несмотря на множество наблюдаемых случаев плавных переходов между видами, гибридных зон, кольцевых видов и тому подобного, большинство видов все-таки дискретны – имеют четкие границы. Обычно между видами есть заметные разрывы (хиатусы) как в геномных последовательностях, так и по видимым признакам. Если виды все время меняются, почему разнообразие все-таки структурировано, почему мы не наблюдаем повсюду одни лишь плавные переходы?
Одна из причин – дискретность экологических условий и пониженная конкурентоспособность промежуточных форм. Например, в тайге условия одни, среди арктических льдов – другие, к первым условиям лучше приспособлен бурый медведь, ко вторым – белый. Промежуточные формы в обоих биотопах будут проигрывать в конкурентной борьбе живущим там специалистам. О таком механизме формирования дискретности, основанном на пониженной приспособленности малочисленных промежуточных форм, писал еще Дарвин. Дополнительное объяснение можно получить из аналогии с языками.
Языки тоже в основном дискретны. Конечно, в зонах смешения разноязычных народов часто формируются гибридные диалекты (аналог гибридных зон в биологии). Но все-таки большинство людей говорит на каком-то определенном языке, а не на смешанном диалекте. Главная причина дискретности
вбиологии и лингвистике, по-видимому, одна и та же. Человеку выгодно уметь свободно и полноценно обмениваться информацией с достаточно большой популяцией себе подобных. Говорить на смеси русского и немецкого неудобно: плохо будут понимать и русские, и немцы. Живым организмам точно так же выгодно уметь свободно и полноценно обмениваться генами (смешивая их в потомстве) с достаточно многочисленной группой особей. В пределах каждого генофонда гены под действием отбора притираются, приспосабливаются друг к другу. Иметь в своем геноме смесь генов, приспособленных к разным генофондам, невыгодно, потому что потомство такого организма от скрещивания с любой из «чистых» форм, скорее всего, будет иметь пониженную жизнеспособность. Преимущество свободного обмена информацией (словесной или генетической) в пределах достаточно больших популяций в тенденции приводит к формированию и поддержанию наблюдаемой дискретности. В природе часто возникают межвидовые гибриды, но численность их, как правило, остается небольшой, а их потомство в будущем может постепенно разойтись по двум исходным видам.
242
Попадая в генофонд вида А, смешанный набор генов гибридной особи будет под действием отбора постепенно очищен от «посторонних примесей», так что в итоге в генофонде останутся только гены, хорошо приспособленные именно к этому генофонду. То же самое произойдет и с теми генами гибрида, которые будут подвергаться отбору в пределах генофонда Б. Именно поэтому межвидовая гибридизация, явление довольно частое, лишь изредка приводит к слиянию двух разошедшихся видов в один.
–––––
Хоть биологи и не могут дать четкого определения вида, видообразование (появление новых видов) является фундаментальным эволюционным процессом. Опять-таки это относится в первую очередь к организмам, размножающимся половым путем. Микробиологи, скажем по секрету, уже махнули рукой на попытки выстроить «нормальную» классификацию своих объектов на видовом уровне и предпочитают говорить о линиях, штаммах и клонах. Считать ли, например, бактерий, выработавших в эксперименте Ленски способность питаться цитратом, новым видом бактерий? Никто не знает. Непонятно, как решать подобные вопросы в принципе. Разве что голосованием.
Но у эукариот, практикующих половое размножение, понятие «вид» – нечто большее, чем просто условность. Пока две популяции не превратились в два разных вида, между ними продолжается генетический обмен, их генофонд развивается как единое целое, их эволюционные судьбы тесно связаны. Они могут выработать мелкие различия, адаптируясь к местным условиям, как хомячки в районах со светлой и темной почвой, но поток генов между популяциями будет эти различия постоянно размывать. Чтобы различия между популяциями начали накапливаться, чтобы популяция получила шанс пойти своим, независимым эволюционным путем, поток генов, связывающий ее с другими популяциями, должен прекратиться или как минимум резко уменьшиться. Должна возникнуть репродуктивная изоляция. Это обязательное условие видообразования (Mayr, 1942). История рождения нового вида – это история последовательного формирования разных типов репродуктивной изоляции, накладывающихся друг на друга. Одни формы изоляции начинают процесс видообразования, другие его продолжают, третьи заканчивают, делая разделение необратимым. Последовательность событий, приводящих к видообразованию, может быть очень разной.
В роли первичного изолирующего фактора может выступать географический барьер (морской пролив, пустыня, горный хребет, широкая река). В этом случае говорят об аллопатрическом или географическом видообразовании. Но барьер может и не быть таким масштабным, он может иметь совсем другую, негеографическую природу. Барьером могут стать экологические или поведенческие свойства видов. Например, популяции могут разойтись по разным кормовым растениям или по срокам размножения. Такое видообразование, происходящее без помощи географического барьера, называют симпатрическим или экологическим.
Независимо от природы первичного изолирующего барьера разделившиеся популяции рано или поздно становятся генетически несовместимыми. Возникает постзиготическая изоляция, проявляющаяся в снижении жизнеспособности или плодовитости гибридного потомства. «Постзиготическая» значит «после образования зиготы». То есть происходит скрещивание, оплодотворение, формируется зигота – а все проблемы начинаются уже потом. Состоят они в пониженной приспособленности этой зиготы и организма, который из нее вырастет. Постзиготическая изоляция – не адаптация, развившаяся под действием отбора. Она лишь побочное следствие накопления генетических различий между популяциями (см.
рис. 36).
243
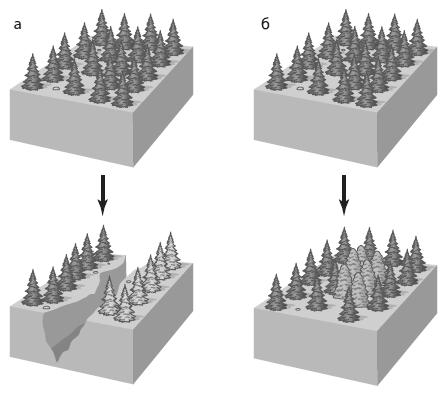
Рис. 36 – Аллопатрическое (a) и симпатрическое (б) видообразование (по А.В. Маркову, 2014)
Если две разобщенные популяции, между которыми уже возникла частичная постзиготическая изоляция, вдруг встретятся (например, из-за исчезновения географического барьера, разделявшего их), то они, скорее всего, поначалу будут охотно скрещиваться друг с другом. Но гибридное потомство будет иметь пониженную приспособленность, а это значит, что отбор поддержит любые мутации, снижающие вероятность гибридизации и способствующие выбору «своих» в качестве брачных партнеров. Этот эффект называется «усилением» (reinforcement), и он приводит к тому, что представители разошедшихся популяций перестают даже пытаться спариться друг с другом. На этом этапе могут быстро развиться разнообразные изолирующие механизмы, в том числе морфологические (например, различия в строении копулятивных органов) и поведенческие (разные брачные наряды и ритуалы ухаживания). Эти механизмы уже являются настоящими адаптациями для предотвращения гибридизации, поскольку они развились под действием отбора. Изоляция, порождаемая такими адаптациями, называется презиготической (до образования зиготы).
Существует несколько возможных сценариев видообразования, в которых разные типы репродуктивной изоляции возникают в разной последовательности и по разным причинам (см. рис. 37). Например, презиготическая изоляция может сформироваться и без помощи «усиления»: либо медленно, вместе с постзиготической, как побочный эффект постепенного накопления различий (морфологических и поведенческих) между географически разобщенными популяциями, либо быстро, например из-за мутации, изменившей сроки цветения или выбор кормового растения. В последнем случае по мере адаптации к новой нише может автоматически развиться постзиготическая изоляция (гибриды будут хуже приспособлены и к новой, и к старой нише, чем родительские формы). Это, в свою очередь, будет способствовать «усилению» презиготической изоляции под действием отбора. Презиготическая изоляция, существующая достаточно долго, может действовать подобно географическому барьеру, приводя к постепенному развитию постзиготической изоляции, и т. д. Итог один: исходный вид разделяется на два репродуктивно изолированных дочерних вида. Мы разберем несколько примеров, показывающих, что в природе могут срабатывать разные сценарии видообразования в зависимости от ситуации.
244
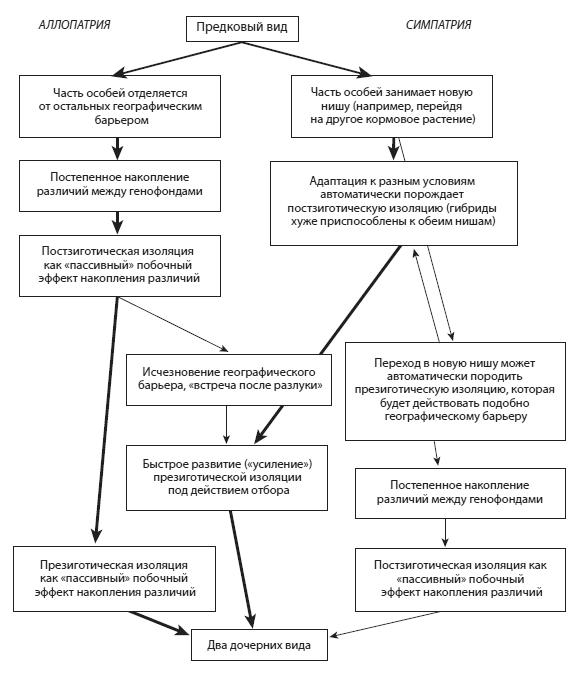
Рис. 37 – Возможные сценарии видообразования. Жирными стрелками выделены последовательности событий, предположительно являющиеся наиболее
типичными (часто встречающимися в природе) (по А.В. Маркову, 2014)
Друг от друга все дальше и дальше
Формирование постзиготической изоляции – своего рода точка невозвращения, после которой у двух популяций остается мало шансов слиться обратно в один вид. Многочисленные факты говорят о том, что постзиготическая изоляция возникает автоматически, сама собой, без помощи какого-либо «отбора на несовместимость» или других специальных механизмов. Единственное, что для этого нужно, – это фактическая разобщенность двух популяций (отсутствие генетического обмена – неважно, по какой причине) и время. Причем времени может потребоваться много: до 10 млн лет и даже
245
больше. Ну а если на популяции действует интенсивный разнонаправленный отбор, постзиготическая изоляция может развиться очень быстро, всего за несколько сотен поколений (на это указывают результаты экспериментов, с которыми мы вскоре познакомимся).
В конце 1930-х – начале 1940-х годов прошлого века Феодосий Добжанский и Герман Мёллер предложили простую модель, согласно которой постзиготическая несовместимость разобщенных популяций развивается как неизбежный побочный эффект их разобщенности. Допустим, в исходной популяции, еще до разделения, все особи имели генотип A1B1C1D1 (предположим для простоты, что организмы гаплоидные). Но вот популяция по какой-то причине разделилась – например, несколько особей занесло бурей на необитаемый остров. После этого в каждой из двух популяций, материковой и островной, будут независимо закрепляться разные мутации – неважно, под действием отбора или дрейфа. Например, в островной популяции закрепится мутация в локусе A. Теперь генотип у обитателей острова будет A2B1C1D1. Может ли это привести к постзиготической несовместимости с материковой популяцией, где сохранился исходный генотип? Вряд ли, потому что ген А2 вполне совместим с генами B1, C1 и D1 (иначе он бы не смог закрепиться в островной популяции). Но вот по прошествии какого-то времени в материковой популяции закрепилась другая мутация, например в локусе B. Теперь генотип материковых особей – A1B2C1D1. Ген В2 совместим с A1, C1 и D1: это было «проверено» отбором, который в противном случае отбраковал бы ген B2 и не дал ему закрепиться в материковой популяции. Однако ген B2 не проверялся отбором на совместимость с геном A2, который закрепился на острове. Следовательно, с некоторой вероятностью эти гены могут оказаться несовместимыми.
Очевидно, что вероятность появления пары несовместимых генов в двух популяциях будет расти пропорционально квадрату числа различающихся генов (т. е. квадрату «генетического расстояния» между популяциями). Например, если в каждой популяции закрепилось по одной мутации, как в нашем примере, то существует только одна пара потенциально несовместимых (не проверявшихся отбором на совместимость) генов: А2 и B2. Если закрепилось по две мутации (например, в островной популяции генотип стал A2B1C2D1, в материковой A1B2C1D2), то таких не проверенных отбором пар будет уже четыре: конфликт может возникнуть между A2 и B2, A2 и D2, C2 и B2, C2 и D2. Если в каждой популяции закрепилось по три мутации (все в разных локусах), число потенциально несовместимых пар будет равно девяти, и т. д.
Таким образом, модель Добжанского – Мёллера предсказывает, что число генов, ответственных за генетическую несовместимость, должно расти с ускорением, как снежный ком. Если предположить, что различия накапливаются с постоянной скоростью (т. е. исходить из допущения о равномерном ходе молекулярных часов), то число «генов несовместимости» должно быть пропорционально квадрату времени, прошедшего с момента разделения популяций.
Модель предполагает, что главную роль в формировании постзиготической изоляции играют попарные антагонистические взаимодействия («антагонистический эпистаз») между разными локусами. Если учитывать возможность конфликтов не только между двумя, но и между тремя и более локусами, то картина существенно усложнится.
В течение 70 с лишним лет, прошедших с момента появления этой модели, в ее работоспособности мало кто сомневался. Ее преподавали в университетах, а генетики, опираясь на нее, занимались поиском конкретных «генов несовместимости», порой весьма успешно. Но вот проверить ее напрямую не удавалось. Для этого нужно сравнить число генетических различий с числом «генов несовместимости» в нескольких парах родственных видов, различающихся по времени своего расхождения. Главная проблема в том, как подсчитать «гены несовместимости».
Лишь в 2010 году сразу двум научным коллективам одновременно удалось преодолеть эту трудность. Первая работа выполнена сотрудниками Чикагского университета на дрозофилах (Matute et al., 2010). Среди ее авторов – Джерри Койн, написавший
246
замечательную научно-популярную книгу Why evolution is true (Coyne, 2009) о доказательствах эволюции.
Ученые воспользовались тем обстоятельством, что некоторые виды дрозофил, хотя и разошлись давно, все еще сохранили способность к гибридизации. Авторы скрещивали
самок Drosophila melanogaster |
с самцами двух других видов: D. simulans и D. santomea. |
Эволюционные пути видов |
melanogaster и simulans разошлись 5,4 млн лет назад, |
melanogaster и santomea – примерно 12,8 млн лет назад. Таким образом, время независимого существования в двух парах видов различается в 2,4 раза, и количество генетических различий тоже (собственно, по последнему и вычислили первое!). Если гипотеза Добжанского – Мёллера верна, число «генов несовместимости» в паре melanogaster – santomea должно быть примерно в 2,42 = 5,76 раза больше, чем в паре melanogaster – simulans.
Чтобы подсчитать «гены несовместимости» в каждой паре, авторы использовали большую, включающую сотни линий, коллекцию дрозофил D. melanogaster с различными делециями – выпадениями небольших участков генома. У мух из этих линий делеции поддерживаются в гетерозиготном состоянии. То есть у каждой мухи в одной хромосоме отсутствует какой-то кусочек, но в другой, парной хромосоме этот кусочек имеется. При этом вторая, целая хромосома, во-первых, «помечена» какой-нибудь доминантной мутацией (чтобы сразу было видно, кому из потомства она досталась), во-вторых, она неспособна рекомбинировать – обмениваться участками с парной хромосомой во время мейоза. Половина половых клеток такой мухи получает целую хромосому с меткой, а другая половина – хромосому с недостающим фрагментом и без метки.
Делеции, имеющихся в разных линиях, в общей сложности покрывают почти весь геном. Авторы скрещивали самок из этих линий (напомним, все они относятся к виду melanogaster) с самцами simulans и santomea. От таких межвидовых скрещиваний рождаются только самки. Потомство мужского пола погибает на ранних стадиях.
Здесь нужно сделать небольшое отступление, чтобы объяснить, почему так получается. Дело в том, что у самцов дрозофил, как и у людей, только одна X-хромосома, а у самок – две. Поэтому гены, расположенные в X-хромосоме, у самцов имеются в единственном экземпляре. Если хоть в одном из них имеется летальная мутация, самец обречен. У самки же есть вторая копия X-хромосомы, в которой данный ген может быть нормальным. Нормальная копия гена «перекрывает» действие испорченной копии, и муха выживает.
Что же за летальные мутации сидят в генах Х-хромосомы гибридных самцов? Это, очевидно, те самые «гены несовместимости», ради поиска которых все и было затеяно. Они конфликтуют с какими-то другими генами в геноме того вида, которому не принадлежит данная X-хромосома. Причем из того факта, что гибридные самки все-таки выживают, можно сделать вывод, что все «гены несовместимости» в данной паре видов рецессивны. Допустим, если несовместимыми являются гены A2 и B2, причем ген B расположен на Х-хромосоме, то гибридные самки с генотипом A1A2B1B2 будут жить, а самцы с генотипом
A1A2B2 погибнут.
Итак, при скрещивании самки melanogaster с самцом simulans или santomea все потомство состоит из дочерей. Причем эти дочери будут двух типов. Первому типу достанется от матери, самки melanogaster, хромосома с отсутствующим кусочком. У таких гибридных мух соответствующий кусочек будет присутствовать в единственном экземпляре
– в парной хромосоме, полученной от отца. Таким образом, отцовские гены, расположенные в этом кусочке, окажутся в «оголенном» (гемизиготном) состоянии – как если бы они сидели в X-хромосоме самца. Если в этом фрагменте хромосомы имеется рецессивный «ген несовместимости», гибридные самки будут иметь пониженную жизнеспособность или вообще погибнут. Второй тип дочерей получит от матери melanogaster целую хромосому с меткой (по метке можно определить, к какому из двух типов относится данная гибридная самка). Все гены будут в двух экземплярах, и рецессивная несовместимость не проявится, т. е. все самки второго типа должны выжить (рис. 38).
247
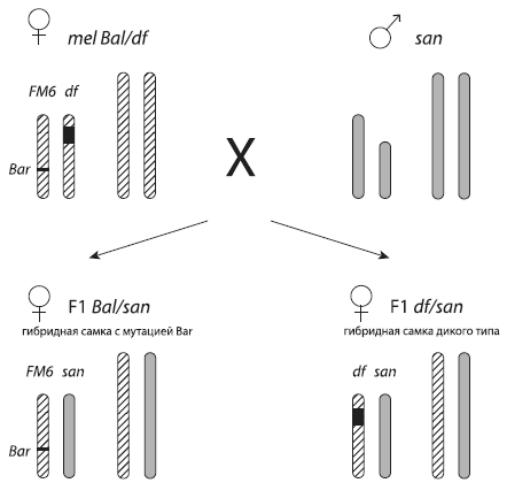
Таким образом, по количественному соотношению гибридных дочерей первого и второго типа можно определить, содержится ли в данном кусочке хромосомы «ген несовместимости». Если обоих типов примерно поровну – значит, «гена несовместимости» там нет; если самок первого типа оказалось существенно меньше, чем второго, значит, он там есть.
Рис. 38 – Схема экспериментов с дрозофилами. Хромосомы melanogaster (mel) показаны косой штриховкой, хромосомы santomea (san) – серым цветом. Скрещивается самка melanogaster с самцом santomea. Самая короткая – Y-хромосома , X-хромосома подлиннее. Черным показан отсутствующий участок хромосомы (в данном случае – X-хромосомы). Хромосома с отсутствующим участком помечена буквами df (от deficiency). Вторая X-хромосома самки melanogaster – целая и помеченная доминантной мутацией Bar. В результате межвидового скрещивания получаются гибридные дочери двух типов: одни несут «целую» материнскую хромосому с меткой, другие получают от матери «урезанную» хромосому df. По выживаемости дочерей второго типа можно понять, есть ли в том кусочке ДНК, который был вырезан из хромосомы df, «ген несовместимости». Из Matute et al., 2010
Таким непростым способом, скрещивая самок из пятисот линий melanogaster с различными делециями с самцами simulans и santomea, авторы просканировали 80 % мушиного генома в поисках «генов несовместимости». Им удалось выявить десять участков, содержащих «гены несовместимости», в паре melanogaster – simulans. В паре melanogaster – santomea таких участков оказалось 65, т. е. в 6,5 раза больше. Можно заметить, что эта величина близка к числу, предсказанному моделью Добжанского – Мёллера (5,74). При помощи сложных статистических методов авторы подтвердили, что их результаты гораздо лучше соответствуют предсказанию модели Добжанского – Мёллера об ускоряющемся, параболическом росте числа «генов несовместимости», чем альтернативной гипотезе о
248
линейном росте. Так классическая модель наконец-то получила красивое подтверждение. Вторая работа была выполнена на растениях рода Solanum (паслен) сотрудниками
Мемфисского и Индианского университетов (Moyle, Nakazato, 2010). Использовалось не три, как в первом исследовании, а четыре близких вида, различающихся по времени расхождения. Здесь тоже авторам понадобилась громадная коллекция подопытных линий, но на этот раз это были линии не мух, а томатов, и различались они не делециями, а небольшими фрагментами геномов диких видов рода Solanum, «встроенными» в геном томата. Получены эти линии были путем межвидовой гибридизации и последующих многократных «возвратных» скрещиваний с помидорами. Для каждой линии было известно, какой именно чужеродный фрагмент внедрился в ее геном. У каждой линии авторы измерили жизнеспособность семян и пыльцы. Если в данной линии оба показателя были в норме, из этого делался вывод, что фрагмент ДНК чужого вида, встроившийся в геном этой линии, не содержит «генов несовместимости». Таким способом удалось подсчитать, как и в первом исследовании, число участков генома, содержащих «гены несовместимости». Расчеты проводились раздельно для жизнеспособности семян и пыльцы.
Результаты по жизнеспособности семян полностью совпали с предсказанием модели Добжанского – Мёллера: число «генов несовместимости», влияющих на этот признак, растет пропорционально квадрату генетической дистанции. С пыльцой, правда, получился отрицательный результат: число «генов несовместимости», влияющих на этот признак, по мере увеличения генетической дистанции растет линейно, а не по параболе. Почему – пока не ясно. Авторы предполагают, что на стерильность пыльцы сильно влияют не попарные, а более сложные взаимодействия между многими локусами.
Так или иначе, обоим научным коллективам удалось получить эмпирические подтверждения адекватности модели Добжанского – Мёллера по крайней мере для некоторых ситуаций. Отрадно, что классики и на этот раз оказались недалеки от истины.
Вид из пробирки
Биологи давно мечтали воспроизвести процесс видообразования в лаборатории. Успехи в этой области достигнуты немалые. Все этапы и «слагаемые» этого процесса, как выяснилось, вполне воспроизводимы в эволюционном эксперименте. И если биологи то и дело не объявляют торжественно об искусственном создании настоящего нового вида, то только из-за расплывчатости критериев вида, о которой говорилось выше. Причем среди применяемых критериев есть и такие, которые просто «по определению» не позволяют назвать новым видом что-либо выведенное в лаборатории (или обособившееся недостаточно давно).
Хотя существует множество «хороших» (признаваемых большинством специалистов) видов, способных скрещиваться друг с другом и давать плодовитое гибридное потомство, главным критерием вида для организмов с половым размножением все-таки считается репродуктивная изоляция. Именно на ней чаще всего и сосредоточиваются усилия биологов, ставящих эксперименты по искусственному видообразованию.
Водних экспериментах изучают зарождение презиготической изоляции. Для этого разные линии подопытных организмов подвергают разнонаправленному отбору (например, помещают в контрастные неблагоприятные условия и выращивают в течение многих поколений), а затем смотрят, не отбило ли у них охоту к перекрестному скрещиванию. Иногда охоту действительно отбивает, животные начинают избирательно скрещиваться со «своими», т. е. теми, чьи предки приспосабливались к таким же условиям (мы еще поговорим об этих опытах).
Вдругих экспериментах изучают формирование постзиготической изоляции, т. е. пониженной приспособленности гибридов. Эти работы показали, что под действием разнонаправленного отбора такая изоляция может сформироваться гораздо быстрее, чем за счет одного лишь случайного накопления различий «по Добжанскому – Мёллеру». Ранее это
249
было показано на тлях (Шапошников, 1965) и дрозофилах (Oliveira, Cordeiro, 1980), а недавно был проведен важный эксперимент на дрожжах (Dettman et al., 2007), с которого мы и начнем.
Дрожжи размножаются как половым путем, так и бесполым – почкованием. Биологи из Университета Торонто (Канада) получили из одной родительской диплоидной клетки 12 подопытных популяций. Изначально все они были генетически идентичны, но потом в результате мутаций и отбора могли накапливать различия. Популяции выращивали в течение 500 поколений в контрастных неблагоприятных условиях: шесть популяций при повышенной солености, шесть – в «голодной» среде с низким содержанием глюкозы. Контрольную (предковую) линию продолжали растить в нормальной среде.
После этого ученые проверили, произошла ли адаптация, т. е. стали ли подопытные популяции лучше приспособлены к своим средам по сравнению с предками. Приспособленность оценивали по скорости почкования клеток в условиях конкуренции с предковой линией. Оказалось, что адаптация произошла во всех 12 линиях, хотя величина приспособленности варьировала. К соленой среде дрожжи приспособились лучше, чем к голодной (вот уж поистине, от голодной диеты скорее умрешь, чем привыкнешь).
Теперь можно было приступить к главной части эксперимента – к выявлению постзиготической изоляции, если таковая возникла. Исследователи получили гибридное потомство, скрестив представителей «соленых» (С) и «голодных» (Г) линий. Кроме того, были получены гибриды подопытных линий с предками (П). Оказалось, что чистые линии С и Г приспособлены к своим средам значительно лучше, чем любые гибриды. Стало быть, частичная постзиготическая изоляция налицо.
Если приспособленность популяций к своим средам складывалась только за счет суммирования эффектов отдельных полезных мутаций, а эпистаза – влияния измененных генов друг на друга – не было, то приспособленность гибридов разного типа должна быть примерно одинаковой. Например, в соленой среде гибриды С/Г и С/П должны демонстрировать одинаковую приспособленность. Это, однако, оказалось не так. Гибриды С/П в соленой среде чувствовали себя лучше, чем С/Г (хотя и хуже, чем чистая линия С). То же самое справедливо и для «голодной» среды: здесь гибриды Г/П оказались более приспособленными, чем С/Г.
Этот результат можно объяснить двояко. Либо адаптации к одной из двух неблагоприятных сред сами по себе снижают приспособленность к альтернативной среде, либо имеет место эпистаз, т. е. гены, ответственные за адаптацию к разным средам, мешают друг другу работать. В первом случае дрожжи из линии С должны расти в голодной среде хуже, чем П, а в соленой среде П должны расти лучше, чем Г. Это не подтвердилось. Следовательно, главную роль в снижении приспособленности гибридов С/Г играет эпистаз.
Но это уже детали. Главный же результат в том, что показана возможность быстрого формирования постзиготической изоляции под действием разнонаправленного отбора. Как мы помним, появление такой изоляции – ключевой этап видообразования.
–––––
Гибридизация не всегда вредна
У американских жаб-лопатоногов виды отделены друг от друга зыбкой гранью. Многие виды нет-нет да и скрещиваются, производя гибридное потомство. Это характерно не только для амфибий: та же картина наблюдается во многих группах млекопитающих, рыб, насекомых. Представление о виде как о совокупности особей, полностью изолированной от всех остальных подобных совокупностей, – абстракция вроде «идеального газа».
Все же конспецифичные (внутривидовые) скрещивания обычно происходят гораздо чаще, чем гетероспецифичные (межвидовые), а потомство от них оказывается более жизнеспособным и плодовитым. Это справедливо и для американских лопатоногов Spea bombifrons и S. multiplicata. Ареалы этих видов частично перекрываются, и в зоне совместного обитания иногда происходит гибридизация. Гибридные жабы отличаются от чистокровных пониженной плодовитостью. Поэтому жабам в общем случае невыгодно скрещиваться с чужаками. Как часто бывает в
250