


Acetylation of the N-terminal domain of H4 (especially at K16)
results in unfolding of 30 nm fiber (due to a reduction of positive charge of the H4 N-terminal domain)
Substitution of H2A by H2A Bbd results in unfolding of 30 nm fiber (because of reduction of negative charge at the surphase of nucleosomal cores)

Ацетилирование гистонов способствует разворачиванию 30 нм хроматиновой фибриллы

Routh et al., (2008) PNAS 105. 8872-8877
При расстоянии между центрами нуклеосом ~ 200 пн присутствие H1 (H5) в стехиометрических количествах имеет ключевое значение для образования 30 нм фибриллы.
При этом олигонуклеосомный фрагмент должен быть достаточно длинным
При расстоянии между центрами нуклеосом ~165 пн ключевую роль в компактизации играют межнуклеосомные взаимодействия
6,1 нуклеосом на 11 нм |
11,2 нуклеосом на 11 нм |

N1

Канонической 30 нм фибриллы просто не существует. В ядре присутствует набор разнообразных компактных
хроматиновых фибрилл, толщина которых близка к 30 нм.


Архитектурные белки хроматина способствуют образованию различных компактных структур
oligonucleosomal array
24 particles 12 particles
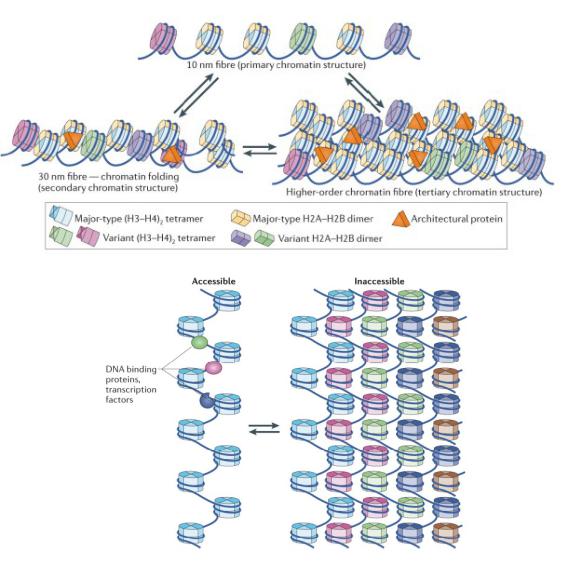

Подвижность нуклеосом в хроматине обеспечивает доступность даже при очень высоком уровне компактизации хроматина (в метафазных хромосомах)