

реципиента, которая осуществляет донашивание и роды. Число успешных результатов такого переноса в последнее время возрастает.
Гаметогенез отличается высокой производительностью. За время половой жизни мужчина продуцирует не менее 500 млрд. сперматозоидов. На пятом месяце эмбриогенеза в зачатке женской половой железы насчитывается 6—7 млн. клетокпредшественниц яйцеклеток. К началу репродуктивного периода в яичниках обнаруживается примерно 100 000 овоцитов. От момента полового созревания до прекращения гаметогенеза в яичниках созревает 400—500 овоцитов.
5.4.ЧЕРЕДОВАНИЕ ГАПЛОИДНОЙ
ИДИПЛОИДНОЙ ФАЗ ЖИЗНЕННОГО ЦИКЛА
Организмы с половым размножением в результате мейоза образуют гаплоидные гаметы. В момент оплодотворения благодаря слиянию двух таких клеток возникает диплоидная зигота. Многократные митотиче-ские деления зиготы и ее потомков приводят к увеличению числа диплоидных клеток, которые строят тело организма в процессе его развития. По достижении последним половой зрелости возобновляется процесс образования гаплоидных гамет. Таким образом, в жизненных циклах организмов, размножающихся половым способом, выделяются две фазы: гаплоидная и диплоидная (рис. 5.11). Относительная продолжительность этих фаз варьирует у представителей различных групп живых существ: у простейших и грибов преобладает первая, у высших растений и животных — вторая.
Удлинение диплофазы в ходе эволюции объясняется преимуществами диплоидного состояния перед гаплоидным. Благодаря гетерозиготности и рецессивности в диплоидном состоянии укрываются от естественного отбора, сохраняются и накапливаются разнообразные аллели. Это повышает объем генетической информации в генофондах популяций и видов, ведет к образованию резерва наследственной изменчивости, от которого зависят эволюционные перспективы. В то же время у гетерозигот вредные рецессивные аллели, не оказывая влияния на развитие фенотипа, не снижают жизнеспособности.
225
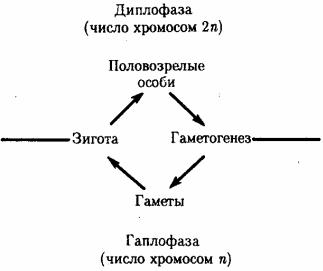
Рис. 5.11. Фазы жизненного цикла многоклеточных животных
5.5. ПУТИ ПРИОБРЕТЕНИЯ ОРГАНИЗМАМИ БИОЛОГИЧЕСКОЙ ИНФОРМАЦИИ
Благодаря генетической рекомбинации, которая закономерно происходит в процессе гаметогенеза и при оплодотворении, половое размножение представляет собой эволюционно обусловленный механизм обмена генетической информацией между организмами одного биологического вида. Некоторые факты из области зоологии и особенно вирусологии и микробиологии указывают и на то, что имеются пути приобретения биологической информации и от организмов других видов (см. 3.6.4.5). Эта информация воспроизводится в фенотипе организма и определяет развитие признаков, не закодированных в генетическом материале родителей. Так, в клетках пищеварительного дивертикула брюхоногого моллюска Elysia viridis сохраняются хлоропласты поедаемой водоросли Codium bragile, в результате чего моллюск приобретает способность к фотосинтезу. Стрекательные капсулы гидроидных полипов, которые поедаются некоторыми реснитчатыми червями, не перевариваются, а перемещаются в эпителиальный пласт и используются червем в качестве орудия защиты. В классической зоологии такие примеры получили название клептогенеза или эволюции путем воровства.
Явление трансдукции заключается в том, что в генетический материал клеткихозяина (бактериальной или эукариотической) встраивается нуклеиновая кислота вируса с фрагментом генома другой клетки. Привносимая таким образом биологическая информация вследствие редупликации чужеродной ДНК может передаваться в ряду клеточных поколений, а также воздействовать на состояние генетической системы клетки-хозяина, изменяя, например, частоту мутирования отдельных генов. Чужеродная ДНК может присутствовать в клетке в виде плазмид и эписом — фрагментов нуклеиновой кислоты, лишенных в отличие от вирусных
226
частиц белковых чехлов. Плазмиды самостоятельны по отношению к хромосомам клетки-хозяина, а эписомы могут встраиваться в них. Биологическая информация плазмид и эписом, проявляясь в фенотипе, дает широкий круг признаков, включая устойчивость к антибиотикам (см. 3.6.3).
Примеры проникновения в организм действующей биологической информации организмов из других таксонов, прежде всего вирусов, описаны у высших животных и человека. Так, сотрудники, длительно работающие в онкологических лабораториях с вирусной опухолью кроликов — папилломой Шопа, имеют, как правило, пониженное содержание в плазме крови аминокислоты аргинина. Объясняется это тем, что вирус папилломы, которым «заражены» такие люди, несет ген синтеза аргиназы, катализирующей обмен аргинина. Будучи активным, этот ген обусловливает образование дополнительных молекул фермента. Описанный факт относится к широкой области так называемого
вирусоносительства или латентных вирусных инфекций. Вирус, присутствуя в клетках организма длительное время и не вызывая собственно патологических изменений, приводит к развитию некоторых фенотипических признаков. У человека известен генетический дефект, который проявляется в серьезных нарушениях развития и зависит от недостатка аргиназы. Открытие способа контролируемого «заражения» таких людей вирусом папилломы Шопа могло бы нормализовать их фенотип.
Технологии геномики (см. разд. 3.2), расшифровывающие нуклеотидные последовательности ДНК, создают почву для целенаправленного изменения или введения в человеческий геном нормальных нуклеотидных последовательностей, что в перспективе может стать основой генотерапии.
Половой процесс служит универсальным механизмом обмена генетической информацией между особями в пределах вида. Рассмотренные факты свидетельствуют о том, что количество и содержание информации, используемой различными организмами для развития и жизнедеятельности, не всегда ограничиваются той, которая была ими приобретена от родителей. Существуют дополнительные пути приобретения биологической информации. Видоспецифическая информация, получаемая в процессе размножения, обеспечивает развитие особей определенного морфофизиологического типа. Дополнительная информация по биологическому содержанию в значительной степени случайна и нередко нарушает реализацию собственной генетической информации хозяина. В связи с этим в эволюции возникли механизмы защиты от проникновения чужеродного наследственного материала. Примером может служить интерферон — белок, вырабатываемый клетками млекопитающих и птиц в ответ на внедрение вирусов и представляющий собой фактор неспецифического противовирусного иммунитета, а также система иммунологического надзора.
227