

При изучении живой материи выде¬ляют несколько уровней ее организации: 1. моле¬кулярный; 2. клеточный; 3. тканевой; 4. органный; 5. онто¬генетический; (или организменный) 6. популяционный; 7. видо¬вой; 8. биогеоценотический; 9. биосферный. Каждому уровню со¬ответствует особая биологическая наука, несколько биологических наук или раздел биологии. На молекулярно-генетическом уровне живые организмы исследуются молекулярной биологией и генетикой; на клеточном – цитологией; на тканевом и органном – анатомией и морфологией, а также физиологией; на онтогенетическом – морфологией и фи¬зиологией; на популяционном – популя-ционной генетикой; на видовом – систе¬матикой и эволюционным учением; на биогеоценотическом – геоботаникой, эко¬логией, биогеоценологией; на биосфер¬ном – биогеоценологией. Медицинская ботаника является фундаментальной медико-биологической дисциплиной и служит базой для освоения профессиональных дисциплин – фармакогнозии и ресурсоведения. Термин «ботаника» происходит от греческого слова ботане , которое значит трава, зелень. Ботаника – наука о растениях, которая изучает закономерности внешнего и внутреннего их строения, особенности жизнедеятельности и географического распространения. Медицинская ботаника – наука о лекарственных растениях и грибах, об их внешнем и внутреннем строении, развитии, жизнедеятельности, биохимическом составе, классификации, распространению, использованию в медицине и прочем Первые датируемые сведения о расте¬ниях содержатся в клинописных таблицах Древнего Востока. Основы ботаники как науки заложили древние греки. Древнегреческий фи¬лософ и естество¬испытатель Теофраст (около 370-285 лет до н. э.) назван К. Линнеем «от¬цом ботаники». После общего упадка есте¬ствознания в средние века ботаника начинает интенсивно развиваться с XVI в. В XVIII – XIX вв. происходит развитие и дифферен¬циация ботаники на отдельные ботанические дисциплины и к первой половине XX в. складывается весь комплекс наук о растениях. Основной раздел ботаники – система¬тика растений. Систематика описывает все ископаемые и современные растительные организмы, разрабатывает классификацию и создает научную основу для изучения филогении растений, то есть выявляет род¬ство таксонов. Морфология исследует осо¬бенности и закономерности внешнего строения растений. Основные успехи в этой области знаний были достигнуты преимущественно в XIX и XX вв. Иссле¬дование внутренней структуры расте¬ний – задача анатомии, которая зароди¬лась в середине XVII в. после изобрете¬ния микроскопа, но, подобно морфоло¬гии, главнейшие открытия были также сделаны в XIX и XX вв. Эмбриология – ботаническая дисцип¬лина, изучающая закономерности образо¬вания и развития зародыша растений. Основы эмбриологии заложены во второй половине XVIII в., но фундамен-тальные открытия были сделаны к нача¬лу XX в. Физиология тесно связана с морфоло¬гией и биохимией растений. Начало фи¬зиологии было положено опытами по пи¬танию растений, осуществленными во второй половине XVIII в. Ныне это ак¬тивно развивающаяся наука, занимаю¬щаяся изучением происходящих в расте¬ниях процессов: фотосинтеза, транспорта веществ, водного обмена, роста, развития, дыхания. География растений зародилась в начале XIX в. Она занимается изуче¬нием основных закономерностей пространственного распространения таксонов (видов, родов и более высоких) и растительных сообществ на Земле. Из бота¬нической географии к концу XIX в. выде¬лилась геоботаника – наука, исследую¬щая основные закономерности формирования, состава, структуры и функционирования растительных сооб¬ществ, а также особенности их пространственного распределения. Экология расте¬ний выясняет отношение растительных организмов к факторам среды и взаимоотношения растений с другими организмами. Она возникла на стыке экологии и ботаники на рубеже XIX и XX вв. и в настоящее время это одна из важнейших отраслей знаний о природе. Помимо фундаментальных ботаниче¬ских дисциплин, выделяют ряд при¬кладных наук, также относимых к бота¬нике. Главнейшей из них считается бота¬ническое ресурсоведение, или экономиче¬ская ботаника. Она рассматривает все аспекты использования растений челове¬ком. В зависимости от объектов и методов их изучения, а также практических по¬требностей выделяют ряд других ботани¬ческих дисциплин. В пределах морфоло¬гии растений выделяют карпологию – раздел знаний о плодах, из анатомии – палинологию, изучающую пыльцу и споры. Пред¬метом исследования палеоботаники яв¬ляются ископаемые растения. У палеобо¬таники свои методы изучения, близкие к методам палеонтологии. Особая роль растений в жизни на Земле состоит в том, что без них было бы не¬возможно существование животных и челове¬ка. Зеленые растения являются основной группой организмов, способных аккумулировать энергию Солнца, создавая органические вещества из не-органических. При этом растения извлекают из атмосферы диоксид углерода (углекислый газ) и выделяют кислород, поддерживая ее по¬стоянный состав. Будучи первичными проду¬центами органических соединений, растения являются определяющим звеном в сложных цепях питания большинства гетеротрофов, на¬селяющих Землю. Благодаря фотосинтезу и непрерывно дей¬ствующей трансформации биогенных элемен¬тов создается устойчивость всей биосферы Зе¬мли и обеспечивается ее нормальное функцио-нирование. Обитая в различных условиях, растения образуют растительные сообщества (фитоценозы), обусловливая разнообразие ландшафтов и экологических ус¬ловий для других организмов. При участии растений формируются по¬чва, торф; скопления ископаемых растений образовали бурый и каменный уголь. Глубо¬кие нарушения растительности неизбежно вле¬кут за собой необратимые изменения биос¬феры и отдельных ее частей и могут оказаться гибельными для человека как биологического вида. Существует пять основных сфер, где прямо или косвенно используются расте¬ния: 1) в качестве продуктов питания для человека и корма для животных, 2) как источники сырья для промышленности и хозяйственной деятельности, 3) как лекарственные средства и сырье для получе¬ния медицинских препаратов, 4) в деко¬ративном озеленении, 5) в охране и улучшении окружающей среды. Пищевое значение растений общеиз¬вестно. В качестве продуктов питания че¬ловека и корма для животных, как прави¬ло, используются части, содержащие за¬пасные питательные вещества или сами вещества, извлеченные тем или иным образом. Потребность в углеводах в ос¬новном удовлетворяется за счет крахмал- и сахаросодержащих растений. Роль источников растительного белка в рацио¬не человека и животных выполняют в ос¬новном некоторые растения из семейства бобовых. Плоды и семена многих видов используют для получения растительных масел. Большинство витаминов и ми¬кроэлементов также поступает вместе со свежей растительной пищей. Существен¬ную роль в питании людей играют пря¬ности и растения, содержащие кофеин – чай и кофе. Техническое использование растений и продуктов из них осуществляется по не¬скольким основным направлениям. На¬иболее широко применяются древесина и волокнистые части растений. Ценность древесины определяется потребностью в ней при изготовлении дере-вянных конструкций любых типов и при производстве бумаги. Сухая перегонка древесины позволяет получить значительное количество важных органи¬ческих веществ, широко упот-ребляемых в промышленности и в быту. Во многих странах древесина – один из основных видов топлива. Остро стоит вопрос о за¬мене угля и нефти энергетически богаты¬ми веществами, продуцируемыми неко¬торыми расте¬ниями. Несмотря на широкое распростране¬ние синтетических волокон, растительные волокна, получаемые из хлопчатника, льна, ко¬нопли, джута, липы сохранили большое зна¬чение при производстве многих тканей. Для лечебных целей растения приме¬няют очень давно. В народных и тради¬ционных медицинах они составляют ос¬новную массу лекарственных средств. В научной медицине России примерно треть препаратов, применяемых для лечения, получают из растений. Считается, что с лечебными целями народы мира ис¬пользуют не менее 21000 видов растений и грибов. В России около 55 видов лекарственных растений культивируется. Подробно с использованием растений в медицине студенты ознакомятся в кур¬сах фармакогнозии и фармакологии. Не менее 1000 видов растений разво¬дят в декоративных целях. Функ¬ционирование всех экологических систем биосферы, частью которой является и че¬ловек, целиком определяется растениями. Растительные ресурсы относятся к категории восполняемых (при правиль¬ной эксплуатации) в противоположность, например, невосполняемым мине-ральным ресурсам. Чаще всего растительные ресурсы делят на ресурсы при¬родной флоры (сюда относятся все дикие виды) и ресурсы культиви¬руемых растений. По объему и значимо¬сти в жизни человечества они существен¬но различаются. Природные ресурсы флоры ограни¬чены и в их естественном объеме могли бы обес¬печить питанием лишь около 10 млн. че¬ловек. Наиболее широко дикорастущие растения исполь¬зуются в качестве технических источни¬ков сырья, в хозяйственной деятельности человека, а также как источник лекарственных средств. Появление культурных растений и возникновение дополни¬тельных растительных ресурсов связано со становлением древнейших человече¬ских цивилизаций. Существование этих цивилизаций могло обеспечиваться толь¬ко определенным «ассортиментом» окультуренных растений, дающих необ¬ходимое количество растительных бел¬ков, жиров и углеводов. Жизнь современ¬ного человека и современная цивилиза¬ция невозможны без широкого ис¬пользования культивируемых растений. Почти все культурные растения (примерно 1500 видов) относятся к покрытосеменным. К середине XX в. культивируемые расте¬ния занимали 15 млн. км2, то есть около 10% всей поверхности суши Земли. Наращивание ресурсов культурных растений возможно в весьма широких пределах как за счет увеличения площа¬дей их возделывания (экстенсификации), так и за счет улучшения агротехники и выведения высокопродуктивных сортов (интенсификация). Считается, что полная мобилизация восполняемых ресурсов, включая растительные, может обеспечить существование на Земле не менее 6 млрд. человек. Около 30% всех выпускаемых меди¬цинских препаратов готовят из лекар¬ственного растительного сырья. Источни¬ком сырья служат как дикорастущие, так и культивируемые растения. Это опреде¬ляет целый комплекс проблем, в которых провизор обязан квалифицированно разо¬браться. Прежде всего, он должен уметь узнавать и характеризовать растения, что делает строго необходимым хорошее знание их морфологии и система¬тики. Подлинность лекарственного расти¬тельного сырья в процессе фармакогностического анализа определяется на основе изучения различных макроскопических и микроскопических признаков. Обяза¬тельным разделом всех стандартов, регу¬лирующих качество лекарственного сы¬рья, является подробная макроскопиче¬ская и микроскопическая характеристики. Макроскопический анализ предполагает хорошее знание морфологии растений и владение соответствующей ботаниче¬ской терминологией. При микроскопиче¬ском анализе провизоры-аналитики изу¬чают растительное сырье анатомически. В этом случае им помогает знание анато¬мии растений. К анатомическим исследо¬ваниям объектов нередко прибегают при судебно-медицинских экспертизах в тех случаях, когда на месте преступления об¬наруживаются растительные остатки. Изучение физиологии растений позво¬ляет понять суть процессов, которые при¬водят к образованию в растениях продук¬тов первичного и вторичного обмена (метаболизма). Многие из них оказы¬ваются фармакологически активными и используются в медицинской прак¬тике. С культивированием лекарственных растений провизор сталкивается относи¬тельно редко, но заготовки дикорастуще¬го лекарственного растительного сырья осуществляются многими аптеками. Поэтому знание флоры региона необходимо для пра¬вильного планирования и организации заготовок. В последние десятилетия различные причины привели к истощению главней¬ших естественных ресурсов ряда лекарственных расте¬ний в традиционных районах заготовок. Актуальными сделались ресурсные иссле¬дования по выявлению новых промыш-ленных массивов лекарственных растений и инвентаризация запасов лекарственного растительного сырья. Эти работы осуществляют провизоры-фармакогносты. Выполнение ресурсных исследований не¬возможно без знания местной флоры, элементов ботанической географии и вла¬дения основными геоботаническими ме¬тодами. Наконец, провизор обязан вы-полнять главнейшие природоохранные мероприятия, которые должны учиты¬ваться при сборе растительного сырья. Это залог длительной эксплуатации за¬рослей дикорастущих лекарственных рас¬тений. Выдающиеся ботаники Украины. Бори́с Никола́евич Аксе́нтьев (1894, Одесса — 19 ноября 1939, Одесса) — ботаник, гидробиолог, фитопатолог, генетик и селекционер растений. Профессор в ряде высших учебных заведений Одессы. Исследовательскую работу начал студентом в 1915 году на Одесской селекционной станции, где был ассистентом по генетике и селекции пшениц у А. А. Сапегина. С 1918 года работал в Одесском университете: до 1921 года был аспирантом Ф. М. Породко на кафедре физиологии растений, затем до 1927 года там же ассистентом, с 1927 по 1930 год — старшим научным сотрудником секции анатомии и физиологии растений научно-исследовательской кафедры биологии. В 1930—1931 годах — старший научный сотрудник зоолого-биологического института; в 1931—1939 годах — в Ботаническом саду Одесского университета. Аксентьев известен вкладом в исследования физиологии растений (проницаемость клеточной плазмы, прорастание семян и их химическая стимуляция), гидробиологии (диатомовые, фитопланктон), фитопатологии (грибные заболевания баклажана), генетики и селекции (пшеница) Дми́трий Константи́нович Зе́ров (укр. Дмитро Костянтинович Зеров, 1895—1971) — украинский ботаник, академик АН УССР. Основные труды относятся к систематике, флористике и филогении споровых растений, болотоведению, ботанической географии, палеоботанике, истории растительности. Главный редактор томов 6, 9—12 двенадцатитомного издания «Флора УССР». Лауреат Государственной премии УССР (1967) за семитомник «Определитель грибов Украины». Михаи́л Васи́льевич Кло́ков (31 июля (12 августа) 1896, Лебедин Харьковской губернии (ныне Сумская область, Украина) — 1981, Киев) — советский ботаник-систематик, исследователь флоры сосудистых растений Украины, доктор биологических наук, профессор, заслуженный деятель науки Украинской ССР, лауреат Сталинской премии и Государственной премии УССР, автор оригинальной фитоэйдологической концепции. Известен также как украинский поэт и литературный критик (под псевдонимом Михаил Доленго). С 1930 по 1941 год заведовал отделом систематики Научно-исследовательского института ботаники Харьковского государственного университета. Одновременно вёл преподавательскую работу на кафедре систематики растений этого же университета. В 1935 году М. В. Клокову присвоено звание профессора, а в 1936 году он получил степень кандидата биологических наук без защиты диссертации. С 1941 по 1944 год находился в эвакуации в городе Кзыл-Орда, где заведовал кафедрой ботаники Объединённого Украинского (Киевского и Харьковского) университета и одновременно занимался изучением местной флоры. С октября 1944 года и до конца жизни М. В. Клоков работал старшим научным сотрудником в отделе систематики Института ботаники имени М. Г. Холодного НАН Украины. В 1947 году М. В. Клоков защитил докторскую диссертацию «Эндемизм украинской флоры». М. В. Клоков был одним из основных авторов и редакторов «Флоры УССР» (с 3-го до 12-го томов) и первого издания «Определителя растений УССР». М. В. Клоков описал с территории СССР 547 новых для науки видов, из них Украины — 389, которые относятся к 99 родам и 30 семействам. Андрей Александрович Потебня (1870—1919) — российский (украинский) миколог, ботаник, специалист в области болезней растений. Значительное внимание Андрей Александрович уделил вопросам анатомии, физиологии и биологии фитопатогенных грибов. Его работы «К вопросу об Exobasidium vitis Pril.» (1897 г.), «Движение плазмы в гифах грибов» (1907 г.), «Новый возбудитель рака яблони Phacidiella discolor (Mont. et Sacc.) A.Pot., его морфология и история развития» (1912 г.), «Новые пиреномицеты лоха (Elaeagnus angustifolia) и сопутствующие конидиальные формы» (1912 г.) и др. внесли значительный вклад в решение частных вопросов микологии и фитопатологии. Андрей Александрович был выдающимся, непревзойденным в украинской микологии коллектором-гербаризатором. Им был собран и упорядочен микологический гербарий, из которого в настоящее время сохранилось свыше 3000 образцов, преимущественно ржавчинных, головневых, сумчатых и анаморфных грибов. Наиболее интересная часть коллекции оформлена в виде 8-томных эксиккатов «Herbarium Rossicum Universitatis Cesareae Charkoviensis». Сюда вошли не только собственные находки А. А. Потебни сборов 1901—1907 гг., но и идентифицированные им материалы В. М. Черняева, А. С. Питра, К. К. Пенго и М. А. Алексеенко. Значительный вклад внес А. А. Потебня в изучение биологического разнообразия грибов Харьковщины. В своих работах «Fungi Imperfecti южной России» (1900 г.) и «Материалы к микологической флоре Курской и Харьковской губернии» (1910 г.) А. А. Потебня отметил на исследованных территориях несколько сотен видов грибов. В 1914 г. Андрей Александрович начал работу над обширной монографией «Грибные паразиты высших растений Харьковской и смежных губерний», которая была призвана обобщить бы сведения о разнообразии фитопатогенных организмов востока Украины. В 1915—1916 гг. вышли в свет первые два тома книги, посвященные бактериям, простейшим, слизевикам, жгутиковым и сумчатым грибам. Преждевременная смерть А. А. Потебни прервала работу над этим уникальным изданием. В фондах Центральной научной библиотеки ХНУ сохраняется незаконченная рукопись третьего тома (посвященного пиреномицетам) и наброски рисунков к ней. Никола́й Григо́рьевич Холо́дный (10 (22) июня 1882, Тамбов — 4 мая 1953, Киев) — учёный-биолог, мыслитель. Работал в области физиологии, анатомии и экологии растений, микробиологии и почвоведения. В области физиологии разработал фитогормональную теорию тропизмов (известна под названием теории Холодного — Вента), объясняющую ростовые движения растений. У Николая Григорьевича Холодного было много последователей и учеников, он оставил более 200 научных работ. Большинство трудов относится к физиологии растений, кроме того ему принадлежат интересные исследования в области экологии растений, микробиологии и почвоведения. Холодный М.Г — основоположник фитогормональной теории тропизмов, объясняющей ростовые движения растений. Им было установлено, что увеличенные дозы ауксина тормозят рост корня или вовсе прекращают его, причем в зоне роста появляется утолщение. Это позволило наметить пути борьбы с сорняками с помощью определённых синтетических веществ. Почти одновременно и независимо от Холодного аналогичные идеи высказал и обосновал экспериментальные данные голландский фитофизиолог Ф. Вент (1928). В мире известна как гипотеза Холодного — Вента. Ещё одним важным вкладом Холодного в науку была разработка проблемы летучих органических соединений атмосферы и выяснение их биологической роли. Холодный Н. Г. проводил опыты по искусственной стимуляции развития растений, которую сейчас широко применяют в сельскохозяйственной практике во всем мире. Большую ценность представляют его исследования по морфологии и физиологии железобактерий. Он также занимался экологическими исследованиями, исследованиями в области почвоведения. Предложил новые методы учета количества почвенных бактерий. Разрабатывал общебиологические вопросы — возникновения жизни на Земле, эволюционной теории, истории науки. Красная книга Украины — официальный государственный документ, который содержит аннотированный перечень редчайших и находящихся под угрозой исчезновения видов животного и растительного мира в границах территории Украины, ее континентального шельфа и морской экономической зоны, а также обобщенные сведения о распространении, современном состоянии этих видов, причинах сокращения численности, и мероприятия по их сохранению и воспроизведению. Занесенные в Красную книгу Украины виды растений и животных подлежат особой охране на всей территории Украины. Организация сохранения видов животных и растений, занесенных в Красную книгу Украины, улучшение среды их обитания или произрастания, создание подходящих условий для размножения в природной среде, разведение и расселение возлагается в границах их компетенции на Кабинет Министров Украины, Советы народных депутатов, городские государственные администрации, исполнительные органы местного самоуправления, Министерство охраны окружающей природной среды Украины и прочие государственные органы, на которые законодательством Украины и Республики Крым возложено осуществление функций в этой сфере. 7 февраля 2002 года парламент Украины принял закон «О Красной книге Украины». Красная книга Украины определяется как основной государственный документ, обобщающий данные о современном состоянии видов животных и растений Украины, которые находятся под угрозой исчезновения, а также меры для их сохранения и восстановления на научно-обоснованных основаниях В 2009 г. вышло третье издание Красной книги Украины. В третье издание Красной книги Украины также внесено 826 видов растений и грибов: сосудистые растения (611 видов), мохообразные (46), водоросли (60), лишайники (52), грибы (57)[2]. Количество видов растений в третьем сравнительно со вторым изданием увеличилось на 285 видов, а во втором сравнительно с первым — на 390 видов. Таким образом, с учетом примерно одинаковых промежутков времени между изданиями Красной книги Украины, наблюдается определенное замедление темпов уменьшения потерь разнообразия видов растений и грибов Украины. В издании Красной книги 2009 года для видов применены следующие категории: • Уязвимый • Исчезающий • Недостаточно известный • Неоцененный • Редкий Уязвимые Виды, которые в ближайшем будущем могут быть отнесены к категории «исчезающих», если продолжится действие факторов, влияющих на их численность. Исчезающие Виды, которые находятся под угрозой исчезновения, сохранение которых является маловероятным, при продолжении губительного действия факторов. Недостаточно известные Виды, которые можно отнести к одной из вышеперечисленных категорий, однако в связи с отсутствием полной достоверной информации, решение данного вопроса остается неопределенным. Редкие Виды с небольшими популяциями, которые в данное время не относятся к категории «исчезающих» или «уязвимых».
Конспект 2
2.1. Строение растительной клетки Клетка - основная форма организации живой материи, элементарная единица организма. Она представляет собой самовоспроизводящуюся систему, которая обособлена от внешней среды и сохраняет определенную концентрацию химических веществ, но одновременно осуществляет постоянный обмен со средой. Клетка - основная структурная единица одноклеточных, колониальных и многоклеточных организмов. Единственная клетка одноклеточного организма универсальна, она выполняет все функции, необходимые для обеспечения жизни и размножения. У многоклеточных организмов клетки чрезвычайно разнообразны по размеру, форме и внутреннему строению. Это разнообразие связано с разделением функций, выполняемых клетками в организме. Несмотря на огромное разнообразие, клетки растений характеризуются общностью строения - это клетки эукариотические, имеющие оформленное ядро. От клеток других эукариот - животных и грибов - их отличают следующие особенности: 1) наличие пластид; 2) наличие клеточной стенки, основным компонентом которой является целлюлоза; 3) хорошо развитая система вакуолей; 4) отсутствие центриолей при делении; 5) рост путем растяжения. Форма и размеры растительных клеток очень разнообразны и зависят от их положения в теле растения и функций, которые они выполняют. Плотно сомкнутые клетки чаще всего имеют форму многогранников, что определяется их взаимным давлением, на срезах они обычно выглядят как 4 – 6-угольники. Клетки, диаметр которых по всем направлениям приблизительно одинаков, называются паренхимными. Прозенхимными называются клетки сильно вытянутые в длину, длина превышает их ширину в 5-6 и более раз. В отличие от клеток животных, взрослые клетки растений всегда имеют постоянную форму, что объясняется присутствием жесткой клеточной стенки. Размеры клеток большинства растений колеблются от 10 до 100 мкм (чаще всего 15-60 мкм), они видны только под микроскопом. Более крупными обычно бывают клетки, запасающие воду и питательные вещества. Мякоть плодов арбуза, лимона, апельсина состоит из столь крупных (несколько миллиметров) клеток, что их можно увидеть невооруженным глазом. Очень большой длины достигают некоторые прозенхимные клетки. Например, лубяные волокна льна имеют длину около40 мм, а крапивы – 80 мм, при этом величина их поперечного сечения остается в микроскопических пределах. Число клеток в растении достигает астрономических величин. Так, один лист дерева насчитывает более 100 млн. клеток. В растительной клетке можно различить три основные части: 1) углеводную клеточную стенку, окружающую клетку снаружи; 2) протопласт – живое содержимое клетки, - прижатый в виде довольно тонкого постенного слоя к клеточной стенке, и 3) вакуоль – пространство в центральной части клетки, заполненное водянистым содержимым – клеточным соком. Клеточная стенка и вакуоль являются продуктами жизнедеятельности протопласта. 2.2. Протопласт Протопласт – активное живое содержимое клетки. Протопласт представляет собой чрезвычайно сложное образование, дифференцированное на различные компоненты, называемые органеллами (органоидами), которые постоянно в нем встречаются, имеют характерное строение и выполняют специфические функции (рис. 2.1 ). К органеллам клетки относятся ядро, пластиды, митохондрии, рибосомы, эндоплазматическая сеть, аппарат Гольджи, лизосомы, микротельца. Органеллы погружены в гиалоплазму, которая обеспечивает их взаимодействие. Гиалоплазма с органеллами, за вычетом ядра, составляет цитоплазму клетки. От клеточной стенки протопласт отделен наружной мембраной – плазмалеммой, от вакуоли - внутренней мембраной – тонопластом. В протопласте осуществляются все основные процессы обмена веществ. Рис. 2.1. Строение растительной клетки по данным электронной микроскопии : 1 – ядро; 2 – ядерная оболочка; 3 – ядерная пора; 4 – ядрышко; 5 – хроматин; 6 – кариоплазма; 7 – клеточная стенка; 8 – плазмалемма; 9 – плазмодесмы; 10 – агранулярная эндоплазматическая сеть; 11 – гранулярная эндоплазматическая сеть; 12 – митохондрия; 13 – рибосомы; 14 – лизосома; 15 – хлоропласт; 16 – диктиосома; 17 – гиалоплазма; 18 – тонопласт; 19 – вакуоль. Химический состав протопласта очень сложен и разнообразен. Каждая клетка характеризуется своим химическим составом в зависимости от физиологических функций. Основными классами конституционных, т. е. входящих в состав протопласта, соединений являются: вода (60-90%), белки (40-50% сухой массы протопласта), нуклеиновые кислоты (1-2%), липиды (2-3%), углеводы и другие органические соединения. В состав протопласта входят и неорганические вещества в виде ионов минеральных солей (2-6%). Белки, нуклеиновые кислоты, липиды и углеводы синтезируются самим протопластом. Помимо конституционных веществ, в клетке присутствуют запасные вещества (временно выключенные из обмена) и отбросы (конечные его продукты). Запасные вещества и отбросы получили обобщенное название эргастических веществ. Эргастические вещества, как правило, накапливаются в клеточном соке вакуолей в растворенном виде или образуют включения – оформленные частицы, видимые в световой микроскоп. К эргастическим обычно относят вещества вторичного синтеза, изучаемые в курсе фармакогнозии, - терпеноиды, алкалоиды, полифенольные соединения. По физическим свойствам протопласт представляет собой многофазный коллоидный раствор (плотность 1,03-1,1). Обычно это гидрозоль, т.е. коллоидная система с преобладанием дисперсионной среды – воды. В живой клетке содержимое протопласта находится в постоянном движении, его можно заметить под микроскопом по передвижению органоидов и включений. Движение может быть вращательным (в одном направлении) или струйчатым (направление токов в разных тяжах цитоплазмы различно). Ток цитоплазмы называется также циклозом. Он обеспечивает лучшую транспортировку веществ и способствует аэрации клетки. Цитоплазма - обязательная часть живой клетки, где происходят все процессы клеточного обмена, кроме синтеза нуклеиновых кислот, совершающегося в ядре. Основу цитоплазмы составляет ее матрикс, или гиалоплазма, в который погружены органеллы. Гиалоплазма – сложная бесцветная, оптически прозрачная коллоидная система, она связывает все погруженные в нее органеллы, обеспечивая их взаимодействие. Гиалоплазма содержит ферменты и активно участвует в клеточном метаболизме, в ней протекают такие биохимические процессы, как гликолиз, синтез аминокислот, синтез жирных кислот и масел и др. Она способна к активному движению и участвует во внутриклеточном транспорте веществ. В основе структуры цитоплазмы лежат биологические мембраны – тончайшие (4-10 нм) пленки, построенные в основном из фосфолипидов и белков – липопротеидов. Молекулы липидов образуют структурную основу мембран. Фосфолипиды располагаются двумя параллельными слоями таким образом, что их гидрофильные части направлены наружу, в водную среду, а гидрофобные остатки жирных кислот – внутрь. Часть молекул белков располагается несплошным слоем на поверхности липидного каркаса с одной или обеих его сторон, часть их погружена в этот каркас, а некоторые проходят через него насквозь, образуя в мембране гидрофильные «поры» (рис. 2.2 ). Большинство мембранных белков представлено различными ферментами. Мембраны – живые компоненты цитоплазмы. Они отграничивают протопласт от внеклеточной среды, создают внешнюю границу органелл и участвуют в создании их внутренней структуры, во многом являясь носителем их функций. Характерной особенностью мембран является их замкнутость, непрерывность – концы их никогда не бывают открытыми. В некоторых особенно активных клетках мембраны могут составлять до 90% сухого вещества цитоплазмы. Одно из основных свойств биологических мембран – их избирательная проницаемость (полупроницаемость): одни вещества проходят через них с трудом или вообще не проходят (барьерное свойство), другие проникают легко. Избирательная проницаемость мембран создает возможность подразделения цитоплазмы на изолированные отсеки – компартменты – различного химического состава, в которых одновременно и независимо друг от друга могут протекать различные биохимические процессы, часто противоположные по направлению. Пограничными мембранами протопласта являются плазмалемма – плазматическая мембрана и тонопласт – вакуолярная мембрана. Плазмалемма – наружная, поверхностная мембрана цитоплазмы, обычно плотно прилегает к клеточной стенке. Она регулирует обмен веществ клетки с окружающей средой, воспринимает раздражения и гормональные стимулы, координирует синтез и сборку целлюлозных микрофибрилл клеточной стенки. Тонопласт регулирует обмен веществ между протопластом и клеточным соком. Рибосомы – маленькие (около 20 нм), почти сферические гранулы, состоящие из рибонуклеопротеидов – комплексов РНК и различных структурных белков. Это единственные органеллы эукариотической клетки, которые не имеют мембран. Рибосомы располагаются в цитоплазме клетки свободно, или же прикрепляются к мембранам эндоплазматической сети. Каждая клетка содержит десятки и сотни тысяч рибосом. Располагаются рибосомы поодиночке либо группами из 4-40 (полирибосомы, или полисомы), где отдельные рибосомы связаны между собой нитевидной молекулой информационной РНК, несущей информацию о структуре белка. Рибосомы (точнее, полисомы) – центры синтеза белка в клетке. ^ Эндоплазматическая сеть (эндоплазматический ретикулум) представляет собой разветвленную трехмерную сеть каналов, пузырьков и цистерн, ограниченных мембранами, пронизывающую гиалоплазму. Эндоплазматическая сеть в клетках, синтезирующих белки, состоит из мембран, несущих на наружной поверхности рибосомы. Такая форма получила название гранулярной, или шероховатой (рис. 2.1 ). Эндоплазматическая сеть, не имеющая рибосом, называется агранулярной, или гладкой. Агранулярная эндоплазматическая сеть принимает участие в синтезе жиров и других липофильных соединений (эфирные масла, смолы, каучук). Эндоплазматическая сеть функционирует как коммуникационная система клетки и используется для транспортировки веществ. Эндоплазматические сети соседних клеток соединяются через цитоплазматические тяжи – плазмодесмы, которые проходят сквозь клеточные стенки. Эндоплазматическая сеть – центр образования и роста клеточных мембран. Она дает начало таким компонентам клетки, как вакуоли, лизосомы, диктиосомы, микротельца. При посредстве эндоплазматической сети осуществляется взаимодействие между органеллами. ^ Аппарат Гольджи назван по имени итальянского ученого К. Гольджи, впервые описавшего его в животных клетках. В клетках растений аппарат Гольджи состоит из отдельных диктиосом, или телец Гольджи и пузырьков Гольджи. Каждая диктиосома представляет собой стопку из 5-7 и более уплощенных округлых цистерн диаметром около 1 мкм, ограниченных мембраной (рис. 2.3). По краям диктиосомы часто переходят в систему тонких ветвящихся трубок. Число диктиосом в клетке сильно колеблется (от 10-50 до нескольких сотен) в зависимости от типа клетки и фазы ее развития. Пузырьки Гольджи различного диаметра отчленяются от краев диктиосомных цистерн или краев трубок и направляются обычно в сторону плазмалеммы или вакуоли. Диктиосомы являются центрами синтеза, накопления и выделения полисахаридов, прежде всего пектиновых веществ и гемицеллюлоз матрикса клеточной стенки и слизей. Пузырьки Гольджи транспортируют полисахариды к плазмалемме. Особенно развит аппарат Гольджи в клетках, интенсивно секретирующих полисахариды. Лизосомы – органеллы, отграниченные от гиалоплазмы мембраной и содержащие гидролитические ферменты, способные разрушать органические соединения. Лизосомы растительных клеток представляют собой мелкие (0,5-2 мкм) цитоплазматические вакуоли и пузырьки – производные эндоплазматической сети или аппарата Гольджи. Основная функция лизосом - локальный автолиз – разрушение отдельных участков цитоплазмы собственной клетки, заканчивающееся образованием на ее месте цитоплазматической вакуоли. Локальный автолиз у растений имеет в первую очередь защитное значение: при временном недостатке питательных веществ клетка может сохранять жизнеспособность за счет переваривания части цитоплазмы. Другая функция лизосом – удаление изношенных или избыточных клеточных органелл, а также очищение полости клетки после отмирания ее протопласта, например при образовании водопроводящих элементов. Микротельца – мелкие (0,5-1,5 мкм) сферические органеллы, окруженные одной мембраной. Внутри находится тонкогранулярный плотный матрикс, состоящий из окислительно-восстановительных ферментов. Наиболее известны из микротелец глиоксисомы и пероксисомы. Глиоксисомы участвуют в превращении жирных масел в сахара, что происходит при прорастании семян. В пероксисомах происходят реакции светового дыхания (фотодыхания), при этом в них окисляются продукты фотосинтеза с образованием аминокислот. Митохондрии - округлые или эллиптические, реже нитевидные органеллы диаметром 0,3-1 мкм, окруженные двумя мембранами. Внутренняя мембрана образует выросты в полость митохондрии – кристы, которые значительно увеличивают ее внутреннюю поверхность. Пространство между кристами заполнено матриксом. В матриксе находятся рибосомы, более мелкие, чем рибосомы гиалоплазмы, и нити собственной ДНК ( рис. 2.4). Рис. 2.4. Схемы строения митохондрии в трехмерном изображении (1) и на срезе (2): ВМ – внутренняя мембрана митохондрии; ДНК – нить митохондриальной ДНК; К – криста; Ма – матрикс; НМ – наружная мембрана митохондрии; Р – митохондриальные рибосомы. Митохондрии называют силовыми станциями клетки. В них осуществляется внутриклеточное дыхание, в результате которого органические соединения расщепляются с высвобождением энергии. Эта энергия идет на синтез АТФ – окислительное фосфорилирование. По мере необходимости энергия, запасенная в АТФ, используется для синтеза различных веществ и в различных физиологических процессах. Число митохондрий в клетке колеблется от нескольких единиц до нескольких сотен, особенно их много в секреторных клетках.. Пластиды – органеллы, характерные только для растений. Различают три типа пластид: 1) хлоропласты (пластиды зеленого цвета); 2) хромопласты (пластиды желтого, оранжевого или красного цвета) и лейкопласты (бесцветные пластиды). Обычно в клетке встречаются пластиды только одного типа. Хлоропласты имеют наибольшее значение, в них протекает фотосинтез. Они содержат зеленый пигмент хлорофилл, придающий растениям зеленый цвет, и пигменты, относящиеся к группе каротиноидов. Каротиноиды имеют окраску от желтой и оранжевой до красной и коричневой, но обычно она маскируется хлорофиллом. Каротиноиды делят на каротины, имеющие оранжевую окраску, и ксантофиллы, имеющие желтую окраску. Это липофильные (жирорастворимые) пигменты, по химической структуре они относятся к терпеноидам. Хлоропласты растений имеют форму двояковыпуклой линзы и размеры 4-7 мкм, они хорошо видны в световой микроскоп. Число хлоропластов в фотосинтезирующих клетках может достигать 40-50. У водорослей роль фотосинтетического аппарата выполняют хроматофоры. Их форма разнообразна: чашевидная (хламидомонада), лентовидная (спирогира), пластинчатая (пиннулярия) и др. Хроматофоры значительно крупнее, число их в клетке – от 1 до 5. Хлоропласты имеют сложное строение. От гиалоплазмы они отграничены двумя мембранами – наружной и внутренней. Внутреннее содержимое называется строма. Внутренняя мембрана формирует внутри хлоропласта сложную, строго упорядоченную систему мембран, имеющих форму плоских пузырьков, называемых тилакоидами. Тилакоиды собраны в стопки - граны, напоминающие столбики монет. Граны связаны между собой тилакоидами стромы (межгранными тилакоидами), проходящими через них насквозь вдоль пластиды (рис. 2.5 ). Рис. 2.5. Схема строения хлоропласта в трехмерном изображении (1) и на срезе (2): Вм – внутренняя мембрана; Гр – грана; ДНК – нить пластидной ДНК; НМ – наружная мембрана; Пг – пластоглобула; Р – рибосомы хлоропласта; С – строма; ТиГ – тилакоид граны; ТиМ – межгранный тилакоид. Хлорофилл и хлоропласты образуются только на свету. Растения, выращенные в темноте, не имеют зеленой окраски и называются этиолированными. Вместо типичных хлоропластов в них образуются измененные пластиды, не имеющие развитой внутренней мембранной системы, - этиопласты. Основная функция хлоропластов – фотосинтез, образование органических веществ из неорганических за счет энергии света. Центральная роль в этом процессе принадлежит хлорофиллу. Он поглощает энергию света и направляет ее на осуществление реакций фотосинтеза. Лейкопласты - мелкие бесцветные пластиды. Они встречаются в основном в клетках органов, скрытых от солнечного света, таких как корни, корневища, клубни, семена. Строение их в общих чертах сходно со строением хлоропластов: оболочка из двух мембран, строма, рибосомы, нити ДНК, пластоглобулы аналогичны таковым хлоропластов. Однако, в отличие от хлоропластов, у лейкопластов слабо развита внутренняя мембранная система. Лейкопласты – это органеллы, связанные с синтезом и накоплением запасных питательных веществ, в первую очередь крахмала, редко белков и липидов. Лейкопласты, накапливающие крахмал, называются амилопластами. Этот крахмал имеет вид зерен, в отличие от ассимиляционного крахмала хлоропластов, он называется запасным, или вторичным. Запасной белок может откладываться в форме кристаллов или аморфных включений в так называемых протеинопластах, жирные масла – в виде пластоглобул в элайопластах. Часто в клетках встречаются лейкопласты, не накапливающие запасные питательные вещества, их роль еще до конца не выяснена. На свету лейкопласты могут превращаться в хлоропласты. Хромопласты - пластиды оранжевого, красного и желтого цвета, который обусловлен пигментами, относящимися к группе каротиноидов. Хромопласты встречаются в клетках лепестков многих растений (ноготки, лютик, одуванчик), зрелых плодов (томат, шиповник, рябина, тыква, арбуз), редко - корнеплодов (морковь), а также в осенних листьях. Значение хромопластов до конца еще не выяснено. Большинство из них представляют собой стареющие пластиды. Они, как правило, развиваются из хлоропластов, при этом в пластидах разрушаются хлорофилл и внутренняя мембранная структура, и накапливаются каротиноиды. Это происходит при созревании плодов и пожелтении листьев осенью. Косвенное биологическое значение хромопластов состоит в том, что они обусловливают яркую окраску цветков и плодов, привлекающую насекомых для перекрестного опыления и других животных для распространения плодов. В хромопласты могут превращаться и лейкопласты. Ядро – основная и обязательная часть эукариотической клетки. Ядро является центром управления обменом веществ клетки, ее ростом и развитием, контролирует деятельность всех других органелл. Ядро хранит генетическую информацию и передает ее дочерним клеткам в процессе клеточного деления. Ядро имеется во всех живых растительных клетках, исключение составляют только зрелые членики ситовидных трубок флоэмы. Клетки с удаленным ядром, как правило, быстро погибают. Ядро – самая крупная органелла, его размер составляет 10-25 мкм. Очень большие ядра у половых клеток (до 500 мкм). Форма ядра чаще сферическая или эллипсоидальная, но в сильно удлиненных клетках может быть линзовидной или веретеновидной. Клетка, как правило, содержит одно ядро. В молодых (меристематических) клетках оно обычно занимает центральное положение. По мере роста центральной вакуоли ядро смещается к клеточной стенке и располагается в постенном слое цитоплазмы. По химическому составу ядро резко отличается от остальных органелл высоким (15-30%) содержанием ДНК – вещества наследственности клетки. В ядре сосредоточено 99% ДНК клетки, она образует с ядерными белками комплексы – дезоксирибонуклеопротеиды. В ядре содержатся также в значительных количествах РНК (в основном иРНК и рРНК) и белки. Структура ядра одинакова у всех эукариотических клеток. В ядре различают хроматин и ядрышко, которые погружены в кариоплазму; от цитоплазмы ядро отделено ядерной оболочкой с порами (рис. 2.1 ). Ядерная оболочка состоит из двух мембран. Наружная мембрана, граничащая с гиалоплазмой, несет прикрепленные рибосомы. Оболочка пронизана довольно крупными порами, благодаря которым обмен между цитоплазмой и ядром значительно облегчен; через поры проходят макромолекулы белка, рибонуклеопротеиды, субъединицы рибосом и др. Наружная ядерная мембрана в некоторых местах объединяется с эндоплазматической сетью. ^ Кариоплазма (нуклеоплазма, или ядерный сок) – основное вещество ядра, служит средой для распределения структурных компонентов – хроматина и ядрышка. В ней содержатся ферменты, свободные нуклеотиды, аминокислоты, иРНК, тРНК, продукты жизнедеятельности хромосом и ядрышка. Ядрышко - плотное, сферическое тельце диаметром 1-3 мкм. Обычно в ядре содержатся 1-2, иногда несколько ядрышек. Ядрышки являются основным носителем РНК ядра, состоят из рибонуклеопротеидов. Функция ядрышек – синтез рРНК и образование субъединиц рибосом. Хроматин - важнейшая часть ядра. Хроматин состоит из молекул ДНК, связанных с белками, - дезоксирибонуклеопротеидов. Во время деления клетки хроматин дифференцируется в хромосомы. Хромосомы представляют собой уплотненные спирализованные нити хроматина, они хорошо различимы в метафазе митоза, когда можно подсчитать число хромосом и рассмотреть их форму. Хроматин и хромосомы обеспечивают хранение наследственной информации, ее удвоение и передачу из клетки в клетку. Число и форма хромосом (кариотип) одинаковы во всех клетках тела организмов одного вида. В ядрах соматических (неполовых) клеток содержится диплоидный (двойной) набор хромосом – 2n. Он образуется в результате слияния двух половых клеток с гаплоидным (одинарным) набором хромосом – n. В диплоидном наборе каждая пара хромосом представлена гомологичными хромосомами, происходящими одна от материнского, а другая от отцовского организма. Половые клетки содержат по одной хромосоме из каждой пары гомологичных хромосом. 2.3. Вакуоли и клеточный сок Вакуоли содержатся почти во всех растительных клетках. Они представляют собой полости в клетке, заполненные водянистым содержимым – клеточным соком. От цитоплазмы клеточный сок изолирован избирательно проницаемой вакуолярной мембраной – тонопластом. Тонопласт выполняет барьерные и транспортные функции. Для большинства зрелых клеток растений характерна крупная центральная вакуоль, занимающая до 70-90% объема клетки. При этом протопласт со всеми органеллами располагается в виде очень тонкого постенного слоя, выстилающего клеточную стенку. В постенном протопласте обычно встречаются мелкие цитоплазматические вакуоли. Иногда ядро располагается в центре клетки в ядерном кармашке цитоплазмы, который связан с постенным слоем тончайшими цитоплазматическими тяжами, пересекающими центральную вакуоль. Клеточный сок представляет собой водный раствор различных веществ, являющихся продуктами жизнедеятельности протопласта, в основном, запасными веществами и отбросами. Реакция клеточного сока обычно слабокислая или нейтральная, реже щелочная. Вещества, входящие в состав клеточного сока, чрезвычайно разнообразны. Это углеводы, белки, органические кислоты и их соли, аминокислоты, минеральные ионы, алкалоиды, гликозиды, танниды, пигменты и другие растворимые в воде соединения. Большинство из них относится к группе эргастических веществ – продуктов метаболизма протопласта, которые могут появляться и исчезать в различные периоды жизни клетки. Многие вещества клеточного сока образуются только в растительных клетках. Кроме функции накопления запасных веществ и отбросов, вакуоли в растительных клетках выполняют еще одну важную функцию – поддержание тургора. Концентрация ионов и сахаров в клеточном соке центральной вакуоли, как правило, выше, чем в клеточной стенке; тонопласт значительно замедляет диффузию из вакуоли этих веществ и в то же время легко проницаем для воды. Поэтому вода будет поступать в вакуоль. Такой однонаправленный процесс диффузии воды через избирательно проницаемую мембрану носит название осмоса. Поступающая в клеточный сок вода оказывает давление на постенный протопласт, а через него и на клеточную стенку, вызывая напряженное, упругое ее состояние, или тургор клетки. Тургор обеспечивает сохранение неодревесневшими органами растения формы и положения в пространстве, а также их сопротивление действию механических факторов. Если клетку поместить в гипертонический раствор какой-нибудь нетоксичной соли или сахара (т. е. в раствор большей концентрации, чем концентрация клеточного сока), то происходит осмотический выход воды из вакуоли. В результате этого ее объем сокращается, эластичный постенный протопласт отходит от клеточной стенки, тургор исчезает, наступает плазмолиз клетки(рис.2.9). Рис. 2.9. Схема плазмолиза : 1 – клетка в состоянии тургора; 2 – начало плазмолиза; 3 – полный плазмолиз. Плазмолиз обычно обратим. При помещении клетки в воду или в гипотонический раствор вода снова энергично поглощается центральной вакуолью, протопласт опять прижимается к клеточной стенке, тургор восстанавливается. Плазмолиз может служить показателем живого состояния клетки, мертвая клетка не плазмолизируется, так как не имеет избирательно проницаемых мембран. Тургорное давление не только поддерживает форму неодревесневших частей растений, оно является также одним из факторов роста клетки, обеспечивая рост клеток растяжением, т. е. за счет поглощения воды и увеличения размера вакуоли. У животных клеток центральная вакуоль отсутствует, их рост происходит главным образом за счет увеличения количества цитоплазмы, поэтому размер животных клеток обычно меньше, чем растительных. 2.4. Включения Образование включений вызвано избыточным накоплением некоторых продуктов обмена веществ в определенных участках клетки – в вакуоли, гиалоплазме, различных органеллах, реже в клеточной стенке. Эти вещества часто выпадают в осадок в аморфном виде или в форме кристаллов – включений. Включения имеют определенную форму и хорошо видны в световой микроскоп. По наличию тех или иных включений, их форме и распределению можно отличить одни виды, роды и семейства растений от других, поэтому они часто служат важным диагностическим признаком при анализе лекарственного растительного сырья. Включения представляют собой либо запасные вещества (временно выведенные из обмена веществ соединения), либо конечные продукты обмена. К первой категории включений относятся крахмальные зерна, липидные капли и отложения белков; ко второй – кристаллы некоторых веществ. Крахмальные зерна – наиболее распространенные включения растительных клеток. Полисахарид крахмал – основной тип запасных питательных веществ растений. Крахмальные зерна образуются в строме пластид. В хлоропластах на свету откладываются зерна ассимиляционного (первичного) крахмала, образующиеся при избытке сахаров – продуктов фотосинтеза.. Ночью, когда фотосинтез не происходит, ассимиляционный крахмал с помощью ферментов гидролизуется до сахаров и транспортируется в другие части растения. Запасной (вторичный) крахмал откладывается в амилопластах клеток различных органов растений (корнях, подземных побегах, семенах) из сахаров, притекающих из фотосинтезирующих клеток. При необходимости запасной крахмал также превращается в сахара. Образование крахмальных зерен начинается в определенных точках стромы пластиды, называемых образовательными центрами. Рост зерна происходит путем последовательного отложения слоев крахмала вокруг образовательного центра. Смежные слои в одном зерне могут иметь различный показатель преломления света, и тогда они видны под микроскопом – слоистые крахмальные зерна. Расположение слоев может быть концентрическим (пшеница) или эксцентрическим (картофель) (рис. 2.10 ). Если в амилопласте имеется один образовательный центр, вокруг которого откладываются слои крахмала, то возникает простое зерно, если два и более – то образуется сложное зерно, состоящее как бы из нескольких простых. Полусложное зерно образуется в том случае, если крахмал сначала откладывается вокруг нескольких точек, а затем, после соприкосновения простых зерен, вокруг них возникают общие слои (рис. 2.10 ). Форма, размер, количество в амилопласте и строение (положение образовательного центра, слоистость, наличие или отсутствие трещин) крахмальных зерен часто специфичны для вида растения (рис. 2.10 ). Обычно крахмальные зерна имеют сферическую, яйцевидную или линзовидную форму, однако у картофеля она неправильная. Наиболее крупные зерна (до 100 мкм) характерны для клеток клубней картофеля, в зерновке пшеницы они двух размеров – мелкие (2-9 мкм) и более крупные (30-45 мкм). Для клеток зерновки кукурузы характерны мелкие зерна (5-30 мкм). Сложные крахмальные зерна у риса, овса, гречихи. Рис. 2.10. Крахмальные зерна различных видов растений : А – картофель; Б – пшеница; В – овес; Г – рис; Д – кукуруза; Е – гречиха; 1 – простое зерно; 2 – сложное зерно; 3 – полусложное зерно. Реактивом на крахмал является раствор йода в растворе калия йодида – реактив Люголя. Он окрашивает крахмальные зерна в сине-фиолетовый цвет. Отложения крахмала широко распространены во всех органах растения, но особенно богаты им семена, подземные побеги (клубни, луковицы, корневища), паренхима проводящих тканей корней и стеблей древесных растений. ^ Липидные капли встречаются практически во всех растительных клетках. Жирные масла накапливаются у огромного количества растений и по своему значению являются второй после крахмала формой запасных питательных веществ. Особенно богаты ими семена и плоды. Семена некоторых растений (подсолнечник, хлопчатник, арахис) могут содержать до 40% масла от массы сухого вещества. Поэтому растительные жиры получают, главным образом, из семян. Липидные капли накапливаются непосредственно в гиалоплазме. Они выглядят как мелкие сферические тела, каждая капля отделена от гиалоплазмы мембраной. Иногда липидные капли называют сферосомами. Реактивом на жирное масло является краситель судан III, липидные капли окрашиваются им в оранжево-красный цвет. ^ Белковые включения в виде разнообразных аморфных или кристаллических отложений образуются в различных органеллах клетки. Запасные белки относятся к категории простых белков – протеинов, в отличие от сложных белков – протеидов, составляющих основу протопласта. В наибольшем количестве они откладываются в запасающей ткани сухих семян в виде алейроновых зерен, или белковых телец Белковые включения можно окрасить реактивом Люголя в золотисто-желтый цвет. ^ Кристаллы кальция оксалата часто встречаются в растительных клетках. Они откладываются только в вакуолях. Форма кристаллов кальция оксалата довольно разнообразна (рис. 2.13) и часто специфична для определенных растений, что используется при диагностике лекарственного растительного сырья. Это могут быть одиночные кристаллы ромбоэдрической, октаэдрической или удлиненной формы (листья белены), друзы – звездчатые сростки кристаллов шаровидной формы (листья спорыша, дурмана, сенны, корни ревеня), рафиды – мелкие игольчатые кристаллы, собранные в пучки (листья ландыша, корневища марены), стилоиды – более крупные, палочковидные кристаллы (листья ландыша) и кристаллический песок – скопления множества мелких одиночных кристаллов (листья красавки). Наиболее часто встречаются друзы. Рис. 2.13. Формы кристаллов кальция оксалата : 1,2 – рафиды (1 – вид сбоку, 2 – вид на поперечном срезе); 3 – друза; 4 – кристаллический песок; 5 – одиночный кристалл. Вдоль волокон в коре или вдоль жилок листьев у ряда растений (кора дуба, корни солодки, листья сенны) встречается кристаллоносная обкладка – расположенные параллельными рядами клетки с одиночными кристаллами кальция оксалата (рис.2.14 ). Рис. 2.14. Жилка с кристаллоносной обкладкой в листе сенны. Подтвердить химическую природу кристаллов кальция оксалата можно действием концентрированных минеральных кислот. Под действием кислоты хлористоводородной кристаллы растворяются. При действии кислоты серной кальция оксалат переходит в нерастворимый кальция сульфат (гипс), образующий многочисленные игольчатые кристаллы. К кристаллическим включениям близки цистолиты. Они чаще всего состоят из кальция карбоната или кремнезема и представляют собой гроздевидные образования, возникающие на выступах клеточной стенки, вдающейся внутрь клетки (рис. 2.15 ). Цистолиты характерны для растений семейств крапивных, тутовых. Значение цистолитов пока не выяснено. 2.5. Клеточная стенка ^ Клеточная стенка (клеточная оболочка) – характерный признак растительной клетки, отличающий ее от клетки животной. Клеточная стенка придает клетке определенную форму. Культивируемые на специальных питательных средах клетки растений, у которых ферментативным путем удаляется стенка, всегда принимают сферическую форму. Клеточная стенка придает клетке прочность и защищает протопласт, она уравновешивает тургорное давление и препятствует, таким образом, разрыву плазмалеммы. Совокупность клеточных стенок образует внутренний скелет, поддерживающий тело растения и придающий ему механическую прочность. Клеточная стенка бесцветна и прозрачна, легко пропускает солнечный свет. Обычно стенки пропитаны водой. По системе клеточных стенок осуществляется транспорт воды и растворенных в ней низкомолекулярных соединений (транспорт по апопласту). Клеточная стенка состоит в основном из полисахаридов, которые можно подразделить на скелетные вещества и вещества матрикса. ^ Скелетным веществом клеточной стенки растений является целлюлоза (клетчатка), Качественную реакцию на целлюлозу проводят с реактивом хлор-цинк-йод, целлюлозная клеточная стенка окрашивается в сине-фиолетовый цвет. У грибов скелетным веществом клеточной стенки является хитин – полисахарид, построенный из остатков глюкозамина. Хитин еще более прочен, чем целлюлоза. Клеточные стенки растений часто подвергаются химическим видоизменениям. Одревеснение, или лигнификация происходит в том случае, если в матриксе откладывается лигнин – полимерное соединение фенольной природы, нерастворимое в воде. Одревесневшая клеточная стенка теряет эластичность, резко повышается ее твердость и прочность на сжатие, снижается проницаемость для воды. Реактивами на лигнин являются: 1) флороглюцин и концентрированная хлористоводородная или серная кислота (одревесневшие стенки приобретают вишнево-красную окраску) и 2) сульфат анилина, под действием которого одревесневшие стенки становятся лимонно-желтыми. Лигнификация характерна для стенок клеток проводящей ткани ксилемы (древесины) и механической ткани склеренхимы. Опробковение, или суберинизация происходит в результате отложения с внутренней стороны клеточной стенки гидрофобных полимеров – суберина и воска. Суберин представляет собой смесь эфиров полимерных жирных кислот. Мономерами воска являются жирные спирты и восковые эфиры. Воск легко извлекается органическими растворителями и быстро плавится, образует кристаллы. Суберин – аморфное соединение, не плавится и не растворяется в органических растворителях. Суберин и воск, образуя чередующиеся параллельные слои, выстилают всю полость клетки с внутренней стороны в виде пленки. Субериновая пленка практически непроницаема для воды и для газов, поэтому после ее образования клетка обычно отмирает. Опробковение характерно для стенок клеток покровной ткани пробки. Реактивом на опробковевшую клеточную стенку является судан III, окраска оранжево-красная. Кутинизации подвергаются наружные стенки клеток покровной ткани эпидермы. ^ Кутин и воск откладываются чередующимися слоями на наружной поверхности клеточной стенки в виде пленки – кутикулы. Кутин представляет собой жироподобное полимерное соединение, близкое по химической природе и свойствам суберину. Кутикула предохраняет растение от излишнего испарения воды с поверхности растения. Окрасить ее можно реактивом судан III в оранжево-красный цвет. Минерализация клеточной стенки происходит вследствие отложения в матриксе большого количества минеральных веществ, чаще всего кремнезема (оксида кремния), реже оксалата и карбоната кальция. Минеральные вещества придают стенке твердость и хрупкость. Отложение кремнезема характерно для клеток эпидермы хвощей, осок и злаков. У простых пор диаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон. Поры в двух смежных клетках, как правило, возникают друг против друга. Эти общие поры имеют вид одного канала, разделенного тонкой перегородкой из срединной пластинки и первичной стенки. Такая совокупность двух пор смежных стенок соседних клеток носит название пары пор и функционирует как одно целое. Разделяющий их канал участок стенки называется замыкающей пленкой поры, или поровой мембраной. В живых клетках замыкающая пленка поры пронизана многочисленными плазмодесмами (рис. 2.18 ). Рис. 2.18. Плазмодесмы под электронным микроскопом (схема ): 1 – на продольном срезе; 2 – на поперечном срезе; Пл – плазмалемма; ЦС – центральный стержень плазмодесмы; ЭР – элемент эндоплазматического ретикулума. Таким образом, протопласты клеток не полностью изолированы друг от друга, а сообщаются по каналам плазмодесм. По ним происходит межклеточный транспорт ионов и мелких молекул, а также передаются гормональные стимулы. Посредством плазмодесм протопласты клеток в растительном организме образуют единое целое, называемое симпластом, а транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
Конспект 3
Многоклеточное семенное растение начинает свое существование в виде морфологически простой одноклеточной зиготы. Из зиготы развивается зародыш и, в конечном итоге, зрелое растение. При этом происходит деление клеток, их увеличение и дифференциация, приводящая к формированию тканей. Ткани у растений возникли в ходе эволюции при переходе к существованию на суше, что потребовало от организмов выполнения дополнительных функций по обеспечению питания, укреплению тела, размножению в воздушной сухой среде и т.д. Настоящей тканью называют группу клеток, имеющих общее происхождение, сходное строение и выполняющих одинаковые функции. Эволюция тканей отмечается уже у высокоорганизованных низших растений(багрянок и бурых водорослей), но полного своего развития они получают у наиболее продвинутых прогрессивных групп растений. У более примитивных низших растений и грибов имеются ложные ткани, для клеток которых характерно индивидуальное происхождение. Ткани растений подразделяют: - по наличию протопласта на живые и мертвые, - по форме клеток на паренхимные и прозенхимные, - по толщине оболочек (толсто- и тонкостенные), - по наличию межклетников (плотные и рыхлые), - по основной функции (образовательные и постоянные). Постоянные ткани делятся на первичные и вторичные. По строению и выполняемым функциям постоянные ткани делятся на защитные(покровные и механические) и обеспечивающие обмен веществ(всасывающие, проводящие, выделительные, основные). Образовательные ткани, или меристемы дают начало постоянным тканям, обеспечивают рост органов. Подразделяются на первичные и вторичные. По местоположению в теле и функциям различают апикальные меристемы(рост в длину) и латеральные, или боковые(рост в толщину). Интеркалярные, или вставочные меристемы расположены в основаниях междоузлий побегов и листьев. Травматические, или раневые меристемы возникают в местах повреждения тела. Покровные ткани. Эпидерма – первичная однослойная (реже многослойная) образующаяся из апикальной меристемы защитная и регуляторно-секреторная ткань, покрывающая побег и корень. В составе эпидермы формируются устьица и трихомы. Перидерма – вторичная комплексная ткань, формирующаяся на стеблях древесных растений к концу первого года жизни, включает в себя феллоген(образовательную ткань), мертвую пробку и живую паренхимную феллодерму. Под устьицами из феллогена образуются чечевички. Корка формируется на стволах деревьев и состоит из нескольких слоев перидерм. Эпиблема – покровно-всасывающая ткань корня. Выделительные ткани. Делят на экзогенные секреторные структуры (железистые волоски, железки, чешуйки, нектарники, гидатоды) и эндогенные (секреторные клетки, или идиобласты, вместилища выделений, млечники). Механические ткани. Придают прочность растениям, их клетки могут быть паренхимными и прозенхимными, но всегда с утолщенной пропитанной разными веществами оболочкой. Колленхима – живая ткань с неравномерно утолщенными целлюлозными оболочками. Склеренхима – мертвая ткань с одревесневшими оболочками, состоящая из клеток склереид и волокон, разнообразие которых обусловлено нахождением в разныхчастях растения. Проводящие ткани. Обеспечивают передвижение веществ по растению. Восходящий ток растворов от корня обеспечивают сосуды, или трахеи – прозенхимные одревесневшие мертвые клетки с перфорациями, а также трахеиды, сообщающиеся между собой окаймленными порами. Нисходящий ток органических веществ обеспечивают ситовидные трубки с сопровождающими их клетками-спутницами. Это живые клетки. Проводящие ткани в комплексе с другими образуют сложные ткани: ксилему (древесину) и флоэму (луб). Ксилема и флоэма часто сопровождают друг друга, формируя сосудисто-волокнистые, или проводящие пучки. Основные ткани, или паренхима. Заполняет пространство между другими тканями. Она подразделяется на ассимиляционную, или хлоренхиму, отличающуюся наличием хлоропластов (столбчатую, или палисадную, губчатую, складчатую), запасающую паренхиму (наличие алейронов, крахмала, жирных масел), водонакапливающую паренхиму (у гигрофитов,суккулентов) и воздухоносную аэренхиму (у гидрофитов, гигрофитов). Индивидуальные признаки главных групп тканей и их использование в микроскопическом анализе. Закономерности размещения тканей в вегетативных органах. Меристемы, или образовательные ткани дают начало постоянным тканям, благодаря способности их клеток к делению. Подразделяются на первичные и вторичные, верхушечные (апикальные), боковые (латеральные), вставочные (интеркалярные) и травматические (раневые). Первичные меристемы (прокамбий и перицикл) – это клетки, способные к делению изначально. К ним относятся апикальные, интеркалярные и частично латеральные меристемы. Клетки, которые постоянно находятся в состоянии деления, называются инициалями. Вторичные (камбий и феллоген) – это клетки, вторично начавшие делиться после периода покоя. К ним относятся латеральные и раневые меристемы. Травматические меристемы образуют особую ткань – каллюс, состоящую из паренхимных клеток, прикрывающую место ранения. Каллюсообразовательная способность используется в садоводстве при размножении растений черенками и прививками, а также для культуры тканей. Клетки апикальных меристем всегда паренхимные, многогранные. Между ними нет межклетников, оболочки тонкие, пектино-целлюлозные, цитоплазма густая, ядро крупное в центре клетки, вакуоли отсутствуют или мелкие, пластид и митохондрий мало. Клетки боковых меристем различны по размерам и форме, обычно подобны по форме клеткам тех постоянных тканей, которые они производят. Из паренхимных клеток происходит паренхима проводящих тканей, из прозенхимных с заостренными концами – проводящие элементы. Камбиальные клетки тонкостенные, имеют крупное ядро с большим количеством ядрышек, лейкопласты, небольшие вакуоли. Клетки феллогена прямоугольной формы. Покровные ткани. Эпидерма (кожица) – первичная однослойная(реже многослойная) ткань, образующаяся из апикальной меристемы. Это сложная ткань, клетки которой отличаются по форме, размерам и функциям (устьица,трихомы). Наружная поверхность клеток покрыта кутикулой(или восковым налетом) разной толщины – гладкой, складчатой или бородавчатой.Клетки эпидермы плотно сомкнуты, боковые стенки часто извилисты, у осевых органов и листьев однодольных эпидермальные клетки обычно вытянутые вдоль оси органа. Часто вся клетка заполнена большой вакуолью с бесцветным или окрашенным клеточным соком. Наружная стенка толстая, внутренние тонкие. В пристенном слое цитоплазмы имеются лейкопласты, иногда хлоропласты. В клетках иногда встречаются кристаллы. Эпидермис бывает пропитан кремнеземом или слизью (некоторые плоды и семена). Устьица – образования для регуляции транспирации и газообмена. Устьице состоит из двух замыкающих клеток бобовидной формы, между которыми находится устьичная щель, которая может расширяться и сужаться. Под щелью располагается крупный межклетник – подустьичная полость. Клетки эпидермы, примыкающие к замыкающим клеткам, часто отличаются от остальных клеток, и тогда их называют побочными, или околоустьичными клетками (рис. 3.6 ). Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат. В зависимости от числа побочных клеток и их расположения относительно устьичной щели выделяют несколько типов устьичного аппарата (рис. 3.7 ). В фармакогнозии типы устьичного аппарата используются для диагностики лекарственного растительного сырья. Величина и строение устьичного аппарата разнообразны и имеют важное диагностическое значение для определения вида растения. Различают несколько типов устьичного аппарата: аномоцитный – побочные клетки не выделяются (у всех высших растений, кроме хвощей), анизоцитный – побочных клеток три, одна отличается размером (крестоцветные, мальвовые), парацитный – побочные клетки параллельны устьичной щели (папоротники, хвощи, бобовые, розовые), диацитный – стенки побочных клеток перпендикулярны устьичной щели (губоцветные, гвоздичные), тетрацитный – побочных клеток 4, расположены симметрично – две полярные, две боковые (однодольные), энциклоцитный – побочные клетки образуют узкое кольцо (папоротники, голосеменные), актиноцитный – побочные клетки радиально расходящиеся (цветковые). Диагностическое значение для определения вида растения и его частей имеет характер расположения устьиц и их плотность. У многих растений эпидерма образует наружные одно- или многоклеточные выросты различной формы – трихомы. Трихомы отличаются крайним разнообразием, оставаясь вместе с тем вполне устойчивыми и типичными для определенных видов, родов и даже семейств. Поэтому признаки трихомов широко используются в систематике растений и в фармакогнозии в качестве диагностических. Трихомы делятся на: 1) кроющие и 2) железистые. Железистые трихомы образуют вещества, которые рассматриваются как выделения. Они будут рассмотрены в разделе, посвященном выделительным тканям. ^ Кроющие трихомы имеют вид простых, разветвленных или звездчатых волосков, одно- или многоклеточных (рис. 3.8 ). Кроющие трихомы могут длительное время оставаться живыми, но чаще они быстро отмирают и заполняются воздухом. Густой слой волосков отражает часть солнечных лучей и уменьшает нагрев, создает затишное пространство около эпидермы, что в совокупности снижает транспирацию. Часто волоски образуют покров только там, где располагаются устьица, например на нижней стороне листьев мать-и-мачехи, багульника. Жесткие, колючие волоски защищают растения от поедания животными, сосочки на лепестках привлекают насекомых. Выросты эпидермальных клеток – трихомы – также имеют важное диагностическое значение при анализе лекарственного растительного сырья. Они очень разнообразны, одноклеточные и многоклеточные, одно- и несколькорядные, делятся на кроющие и железистые(элементы секреторных тканей). От трихомов, образующихся только из эпидермальных клеток, следует отличать эмергенцы, в формировании которых принимают участие и более глубоко расположенные ткани. К ним относят шипы розы, малины, ежевики, покрывающие черешки листьев и молодые побеги. Перидерма – вторичная комплексная ткань, образующаяся благодаря активности феллогена. Клетки паренхимные, у пробки – таблитчатые, толстостенные, пропитанные суберином. Несколько слоёв перидермы образуют корку – третичную покровную ткань; ткани, оказавшиеся между слоями перидермы, отмирают. Эпиблема – покровно-всасывающая ткань корня, состоящая из паренхимных клеток, имеющих выросты – корневые волоски. Выделительные ткани. Делят на: 1. Экзогенные секреторные структуры. - железистые волоски и желёзки (многоклеточные на ножке или сидячие структуры, содержащие обычно эфирные масла) - нектарники и осмофоры (многоклеточные видоспецифические структуры на цветках, выделяющие нектар) - гидатоды (структуры на краях листьев у выходов жилок для выделения капель растворов) 2. Эндогенные секреторные структуры - секреторные клетки, или идиобласты (в них накапливаются бальзамы, смолы, камеди, танины, кристаллы, каучук, слизи) - вместилища выделений (схизогенные – с четкими внутренними границами каналы и полости, выстланные секреторными клетками, и лизигенные, образованные в результате лизиса клеток полости с секретом) - млечники (прозенхимные клетки и системы клеток – членистые млечники, содержащие латекс с алкалоидами, гликозидами, смолами и др., млечники бывают неветвистые и ветвистые). Механические ткани. Колленхима – живая с неравномерно утолщенными целлюлозными оболочками ткань из прозенхимных клеток со скошенными концами. Фотосинтезирующая. Расположена в стеблях кольцом или пучками, вдоль жилок и по краям листьев. Различают колленхиму по характеру утолщений стенок на поперечном разрезе: уголковую, пластинчатую, рыхлую. У рыхлой есть межклетники. Склеренхима – мёртвая, с одревесневшими оболочками ткань, по прочности оболочки близки к стали. Состоит из клеток склереид и волокон, разнообразие которых обусловлено нахождением в разных частях растения. Склереиды возникают из основной паренхимы вследствие одревеснения оболочек. Они противостоят сдавливанию. Их клетки очень разнообразны по форме и строению оболочек, что имеет диагностическое значение. Различают брахисклереиды – каменистые паренхимные клетки мякоти плодов, коры, флоэмы и сердцевины; макросклереиды – прозенхимные клетки кожуры семян; остеосклереиды, имеющие форму гантели, в мезофиле листьев; астросклереиды с радиальными отростками, трихосклереиды в виде тонких волосков; волокнистые склереиды в осевых органах. Склеренхимные волокна разбросаны поодиночке или чаще в виде тяжей, цилиндров или обкладок. Они не дают органам разрываться при натяжении. Они разделяются на: - лубяные (флоэмные) – клетки 4-350мм длины с полосато-слоистыми оболочками, косыми порами, частично или полностью одревесневшие, концы клеток различной формы (видоспецифичны), обеспечивающей их крепкое сочленение; - древесинные (либриформ) – клетки короче лубяных, оболочки одревесневшие; - коровые, образующиеся под эпидермой; - перициклические, образующиеся по периферии центрального цилиндра; - обкладочные, формирующиеся вокруг проводящих пучков. Проводящие ткани. Комплексные, состоят из разных элементов. Восходящий ток растворов от корня обеспечивают сосуды, или трахеи – прозенхимные одревесневшие мёртвые клетки с перфорациями, а также трахеиды, сообщающиеся между собой окаймленными порами. Вместе с механическими и паренхимными элементами они образуют ксилему. Нисходящий ток органических веществ обеспечивают ситовидные трубки с сопровождающими их клетками-спутницами. Вместе с механическими и паренхимными тканями они образуют флоэму. Тахеиды – мёртвые прозенхимные клетки, суженные на концах, длиною до 4 мм. Стенки одревесневшие, сообщаются простыми или окаймленными порами. Утолщения стенок лестничные или спиральные.У хвощей, папоротников, плаунов и голосеменных – это единственные проводящие элементы. Трахеи, или сосуды встречаются только у покрытосеменных. Они представляют собой членистые капилляры, клетки которых соединяются через перфорации. Стенки одревесневшие с различными типами утолщений: кольчатые, лестничные, точечные, спиральные, сетчатые. Форма клеток сосудов, характер утолщений, тип поровости и перфораций – это диагностические признаки. Трахеальные элементы в ксилеме располагаются по-разному: кольцесосудистая и рассеяннососудистая древесина. В ксилеме также находятся сердцевинные лучи – ряды тонкостенных паренхимных клеток, идущих от сердцевины к коре в радиальном направлении, а также участки древесинной паренхимы, осуществляющие проводящую и запасающую функции. Часто ксилему сопровождают тяжи или обкладки механической ткани. Ситовидные трубки – элементы флоэмы, имеют прозенхимную форму и сочленяются стенками с многочисленными перфорациями, имеющими вид ситечек – ситовидные поля. Боковые оболочки ситовидных трубок равномерно утолщенные, не одревесневающие. В центре клетки имеется полость, заполненная флоэмным соком, а цитоплазма расположена тонким постенным слоем. Ядра нет, есть митохондрии, лейкопласты и эндоплазматическая сеть. Рядом с ситовидной трубкой находится одна или несколько полноценных клеток-спутниц, обеспечивающих жизнедеятельность цитоплазмы ситовидных трубок, которые функционируют 1-4 года, а потом отмирают вследствие закупорки каллозой. В составе флоэмы также проходят сердцевинные лучи, имеется лубяная паренхима и механические элементы. Ксилема и флоэма часто сопровождают друг друга, формируя сосудисто-волокнистые, или проводящие пучки. Различают закрытые пучки, образованные прокамбием и неспособные к дальнейшему увеличению, и открытые, внутри которых сохраняется камбий, откладывающий флоэму и ксилему длительное время. Различают типы пучков в зависимости от расположения ксилемы и флоэмы: - коллатеральные – ксилема и флоэма расположены рядом на одном радиусе (в осевых органах – наружу флоэма, в листьях - ксилема), закрытые у однодольных, открытые у двудольных; - биколлатеральные – открытые с двумя участками флоэмы кнаружи и вовнутрь (пасленовые, тыквенные, вьюнковые); - концентрические закрытые центрофлоэмные (однодольные) и центроксилемные (папоротниковидные); -радиальные закрытые – флоэма и ксилема чередуются по радиусам (зоны всасывания и проведения корней). Основные ткани, или паренхима, или выполняющая ткань. Заполняет пространство между другими тканями. Подразделяется на: - ассимиляционную (хлоренхима), отличающуюся наличием хлоропластов(столбчатую, или палисадную – состоящую из вытянутых и расположенных перпендикулярно поверхности клеток, губчатую – разнообразные по форме клетки с межклетниками, складчатую – с извилистыми стенками клеток); - запасающую (алейрон, крахмал, жирное масло), находящуюся в сердцевине вегетативных органов, древесине, коре, семенах и плодах, корневищах и корнях; - водонакапливающую(гигрофиты, суккуленты, галофиты) – крупноклеточную тонкостенную с крупными вакуолями ткань; - воздухоносную, или аэренхиму (гидрофиты, гигрофиты) – проветривающую ткань с крупнымимежклетниками
Конспект 4
Тема 4. Вегетативні органи рослин
ЛЕКЦІЯ
Корінь як орган рослини. Функції коренів, види, типи кореневих систем. Зони кореня. Спеціалізація та метаморфози коренів.
Пагін як орган рослини. Функції, частини пагону. Різноманітність стебел за положенням у просторі, формою поперечного перерізу тощо. Визначення бруньки та її будова, класифікація за місцем розташування, структурою, функціями. Метаморфози стебел.
Листок як орган рослини. Частини листків, особливості їх будови та функції. Типи листків та їх морфологічна різноманітність. Розміщення листків. Метаморфози листка.
Значення й використання вегетативних органів рослин.
Корень – основной вегетативный орган растения, выполняющий в типичном случае функцию почвенного питания. Корень – осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы. От побега он морфологически отличается тем, что на нем никогда не образуются листья, а апикальная меристема всегда прикрыта корневым чехликом. Кроме главной функции поглощения веществ из почвы, корни выполняют и другие функции: 1) корни укрепляют («заякоривают») растения в почве, делают возможным вертикальный рост и вынесение побегов наверх; 2) в корнях синтезируются различные вещества, которые затем передвигаются в другие органы растения; 3) в корнях могут откладываться запасные вещества; 4) корни взаимодействуют с корнями других растений, микроорганизмами, грибами, обитающими в почве. Совокупность корней одной особи образует единую в морфологическом и физиологическом отношении корневую систему. В состав корневых систем входят корни различной морфологической природы – главный корень, боковые и придаточные корни. Главный корень развивается из зародышевого корешка. Боковые корни образуются на корне (главном, боковом, придаточном), который по отношению к ним обозначается как материнский. Они возникают на некотором расстоянии от апекса, в направлении от основания корня к его верхушке. Боковые корни закладываются эндогенно, т.е. во внутренних тканях материнского корня. Если бы ветвление происходило в самом апексе, это бы затруднило продвижение корня в почве. Придаточные корни могут возникать и на стеблях, и на листьях, и на корнях.
По
происхождению выделяют следующие типы
корневых систем :
1)
система
главного корня(стержневая)
представлена главным корнем (первого
порядка) с боковыми корнями второго и
последующих порядков (у многих кустарников
и деревьев, большинства двудольных
растений);
2)
система придаточных корней(мочковатая)
развивается
на стеблях, листьях; встречается у
большинства однодольных растений и
многих двудольных, размножающихся
вегетативно;
3)
смешанная корневая система
образована главным и придаточными
корнями с их боковыми ответвлениями
(многие травянистые двудольные).
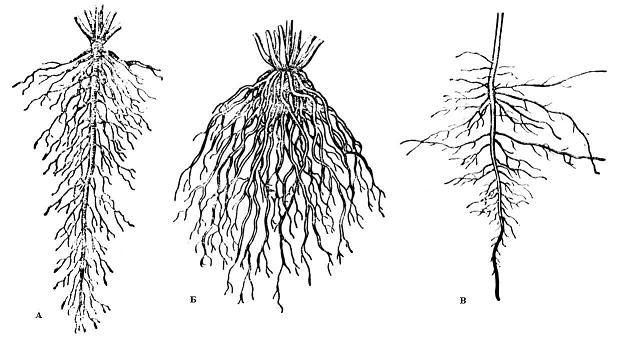 Рис.
4.1. Типы
корневых систем
: А – система главного корня; Б – система
придаточных корней; В – смешанная
корневая система (А и В – стержневые
корневые системы; Б – мочковатая корневая
система).
По форме различают
стержневую
и мочковатую
корневые системы.
В стержневой
корневой
системе главный корень сильно развит
и хорошо заметен среди остальных корней.
В мочковатой
корневой системе главный корень незаметен
или его нет, а корневая система составлена
многочисленными придаточными корнями
( рис. 4.1).
Рис.
4.1. Типы
корневых систем
: А – система главного корня; Б – система
придаточных корней; В – смешанная
корневая система (А и В – стержневые
корневые системы; Б – мочковатая корневая
система).
По форме различают
стержневую
и мочковатую
корневые системы.
В стержневой
корневой
системе главный корень сильно развит
и хорошо заметен среди остальных корней.
В мочковатой
корневой системе главный корень незаметен
или его нет, а корневая система составлена
многочисленными придаточными корнями
( рис. 4.1).
Кончик
корня снаружи всегда прикрыт корневым
чехликом,
защищающим апикальную меристему. Чехлик
состоит из живых клеток и постоянно
обновляется: по мере того, как с его
поверхности слущиваются старые клетки,
на смену им, изнутри, апикальная меристема
образует новые молодые клетки. Наружные
клетки корневого чехлика отслаиваются
еще будучи живыми, они продуцируют
обильную слизь, которая облегчает
продвижение корня среди твердых частиц
почвы. В клетках центральной части
чехлика содержится много крахмальных
зерен. По-видимому, эти зерна служат
статолитами,
т. е. способны перемещаться в клетке при
изменении положения кончика корня в
пространстве, благодаря чему корень
растет всегда в сторону действия силы
тяжести (положительный
геотропизм).
Под
чехликом находится зона
деления,
представленная апикальной меристемой,
в результате деятельности которой
формируются все прочие зоны и ткани
корня. Зона деления имеет размеры около
1 мм. Клетки апикальной меристемы
относительно мелкие, многогранные, с
густой цитоплазмой и крупным ядром.
Вслед
за зоной деления располагается зона
растяжения,
или зона
роста. В этой
зоне клетки почти не делятся, а сильно
растягиваются (растут) в продольном
направлении, вдоль оси корня. Объем
клеток увеличивается за счет поглощения
воды и образования крупных вакуолей,
при этом высокое тургорное давление
проталкивает растущий корень между
частицами почвы. Протяженность зоны
растяжения обычно невелика и не превышает
нескольких миллиметров.
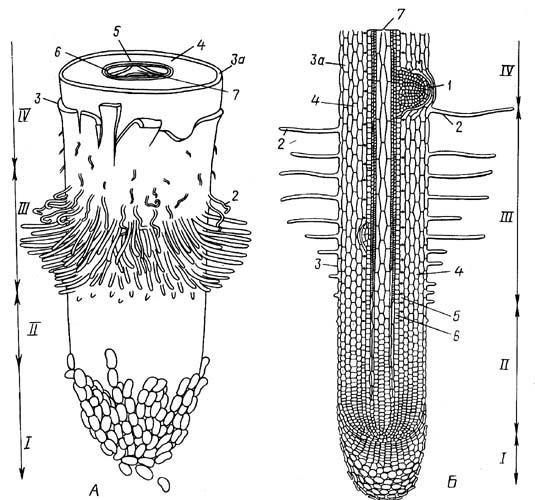 Рис.
4.2. Общий вид
(А) и продольный срез (Б) корневого
окончания (схема
): I – корневой чехлик; II – зоны деления
и растяжения; III – зона всасывания; IV –
начало зоны проведения: 1 – растущий
боковой корень; 2 – корневые волоски; 3
– ризодерма; 3а – экзодерма; 4 – первичная
кора; 5 – эндодерма; 6 – перицикл; 7 –
осевой цилиндр.
Далее идет
зона поглощения,
или зона
всасывания.
В этой зоне покровной тканью является
ризодерма
(эпиблема),
клетки которой несут многочисленные
корневые
волоски.
Растяжение корня прекращается, корневые
волоски плотно охватывают частицы почвы
и как бы срастаются с ними, поглощая
воду и растворенные в ней минеральные
соли. Зона поглощения имеет протяжение
до нескольких сантиметров. Эту зону
называют также зоной
дифференциации,
поскольку именно здесь происходит
образование постоянных первичных
тканей.
Продолжительность жизни
корневого волоска не превышает 10-20 дней.
Выше зоны всасывания, там, где исчезают
корневые волоски, начинается зона
проведения.
По этой части корня вода и растворы
солей, поглощенные корневыми волосками,
транспортируются в вышележащие органы
растения. В зоне проведения формируются
боковые корни (рис.
4.2).
Рис.
4.2. Общий вид
(А) и продольный срез (Б) корневого
окончания (схема
): I – корневой чехлик; II – зоны деления
и растяжения; III – зона всасывания; IV –
начало зоны проведения: 1 – растущий
боковой корень; 2 – корневые волоски; 3
– ризодерма; 3а – экзодерма; 4 – первичная
кора; 5 – эндодерма; 6 – перицикл; 7 –
осевой цилиндр.
Далее идет
зона поглощения,
или зона
всасывания.
В этой зоне покровной тканью является
ризодерма
(эпиблема),
клетки которой несут многочисленные
корневые
волоски.
Растяжение корня прекращается, корневые
волоски плотно охватывают частицы почвы
и как бы срастаются с ними, поглощая
воду и растворенные в ней минеральные
соли. Зона поглощения имеет протяжение
до нескольких сантиметров. Эту зону
называют также зоной
дифференциации,
поскольку именно здесь происходит
образование постоянных первичных
тканей.
Продолжительность жизни
корневого волоска не превышает 10-20 дней.
Выше зоны всасывания, там, где исчезают
корневые волоски, начинается зона
проведения.
По этой части корня вода и растворы
солей, поглощенные корневыми волосками,
транспортируются в вышележащие органы
растения. В зоне проведения формируются
боковые корни (рис.
4.2).
Специализация и метаморфозы корней. У большинства растений в одной и той же корневой системе отчетливо различаются ростовые и сосущие окончания. Ростовые окончания обычно более мощные, быстро удлиняются и продвигаются вглубь почвы. Зона растяжения у них хорошо выражена, и апикальные меристемы работают энергично. Сосущие окончания, возникающие в большом количестве на ростовых корнях, удлиняются медленно, и их апикальные меристемы почти перестают работать. Сосущие окончания как бы останавливаются в почве и интенсивно ее «обсасывают». У древесных растений различают толстые скелетные и полускелетные корни, на которых образуются недолговечные корневые мочки. В состав корневых мочек, непрерывно заменяющих друг друга, входят ростовые и сосущие окончания. Если корни выполняют особые функции, их строение меняется. Резкое, наследственно закрепленное видоизменение органа, вызванное сменой функций, носит название метаморфоза. Видоизменения корней очень разнообразны. Корни многих растений образуют симбиоз с гифами почвенных грибов, называемый микоризой («грибокорень»). Микориза образуется на сосущих корнях в зоне поглощения. Грибной компонент облегчает корням получение воды и минеральных элементов из почвы, часто гифы грибов заменяют корневые волоски. В свою очередь, гриб получает от растения углеводы и другие питательные вещества. Различают два основных типа микоризы. Гифы эктотрофной микоризы образуют чехол, окутывающий корень снаружи. Эктомикориза широко распространена у деревьев и кустарников. Эндотрофная микориза встречается в основном у травянистых растений. Эндомикориза находится внутри корня, гифы внедряются в клетки коровой паренхимы. Микотрофное питание очень широко распространено. Некоторые растения, например орхидные, вообще не могут существовать без симбиоза с грибами. На корнях бобовых возникают особые образования – клубеньки, в которых поселяются бактерии из рода Rhizobium. Эти микроорганизмы способны усваивать атмосферный молекулярный азот, переводя его в связанное состояние. Часть веществ, синтезированных в клубеньках, усваивают растения, бактерии, в свою очередь, используют вещества, находящиеся в корнях. Этот симбиоз имеет большое значение для сельского хозяйства. Бобовые растения благодаря дополнительному источнику азота богаты белками. Они дают ценные пищевые и кормовые продукты и обогащают почву азотистыми веществами. Очень широко распространены запасающие корни. Они обычно утолщены и сильно паренхиматизированы. Сильно утолщенные придаточные корни называют корневыми шишками, или корнеклубнями (георгин, некоторые орхидные). У многих, чаще двулетних, растений со стержневой корневой системой возникает образование, носящее название корнеплода. В образовании корнеплода принимают участие и главный корень, и нижняя часть стебля. У моркови почти весь корнеплод составлен корнем, у репы корень образует лишь самую нижнюю часть корнеплода ( рис. 4.12). Корнеплоды культурных растений возникли в результате длительного отбора. В корнеплодах сильно развита запасающая паренхима и исчезли механические ткани. У моркови, петрушки и других зонтичных паренхима сильно развита во флоэме; у репы, редьки и других крестоцветных – в ксилеме. У свеклы запасные вещества откладываются в паренхиме, образованной деятельностью нескольких добавочных слоев камбия (рис. 4.12 ). У многих луковичных и корневищных растений образуются втягивающие, или контрактильные корни (рис. 4.13, 1 ). Они могут укорачиваться и втягивать побег в почву на оптимальную глубину на время летней засухи или зимних морозов. Втягивающие корни имеют утолщенные основания с поперечной морщинистостью. Воздушные корни образуются у многих тропических эпифитных однодольных из семейств орхидных, ароидных, бромелиевых (рис. 4.13, 3). Эпифиты поселяются на других растениях, но не паразитируют на них, а используют как подпорку для поднятия вверх, к свету. Воздушные корни свободно висят в воздухе и приспособлены к поглощению атмосферной влаги. На их поверхности образуется веламен (рис. 4.14). Веламен, как и ризодерма, образуется из протодермы, но это многослойная ткань. Клетки веламена отмирают, их стенки имеют сетчатые или спиральные утолщения. Через поры и сквозные отверстия в клеточных стенках влага проникает капиллярным путем. Изнутри веламен подстилается экзодермой со сложно построенными пропускными клетками, через которые вода передается в клетки коры и осевого цилиндра (рис. 4.14 ). Веламен встречается и у некоторых наземных однодольных (кливия, хлорофитум), в этом случае он выполняет функцию механической защиты и предохраняет растение от потери воды из первичной коры.
Дыхательные корни, или пневматофоры (рис. 4.13, 2 ) образуются у некоторых тропических древесных растений, живущих в условиях недостатка кислорода (таксодиум, или болотный кипарис; растения мангровых зарослей, обитающие по болотистым берегам океанических побережий). Пневматофоры растут вертикально вверх и высовываются над поверхностью почвы. Через систему отверстий в этих корнях, связанных с аэренхимой, воздух поступает в подводные органы. У некоторых растений для поддержания побегов в воздушной среде образуются дополнительные опорные корни. Они отходят от горизонтальных ветвей кроны и, достигнув поверхности почвы, интенсивно ветвятся, превращаясь в столбовидные образования, поддерживающие крону дерева (столбовидные корни баньяна) (рис. 4.15, 2). Ходульные корни отходят от нижних участков стебля, придавая стеблю устойчивость. Они образуются у растений мангровых зарослей, растительных сообществ, развивающихся на затопляемых во время прилива тропических берегах океанов (рис. 4.15, 3), а также у кукурузы (рис. 4.15, 1 ). У фикуса каучуконосного образуются досковидные корни. В отличие от столбовидных и ходульных, они являются по происхождению не придаточными, а боковыми корнями.
Побег и системы побегов
Побег, как и корень, - основной орган растения. Вегетативные побеги в типичном случае выполняют функцию воздушного питания, но имеют ряд других функций и способны к разнообразным метаморфозам. Спороносные побеги (в том числе и цветок) специализированы как органы репродуктивные, обеспечивающие размножение. Побег образуется верхушечной меристемой как единое целое и, поэтому, представляет собой единый орган того же ранга, что и корень. Однако по сравнению с корнем побег имеет более сложное строение. Вегетативный побег состоит из осевой части – стебля, имеющего цилиндрическую форму, и листьев – плоских боковых органов, сидящих на стебле. Кроме того, обязательной частью побега являются почки – зачатки новых побегов, обеспечивающие нарастание побега и его ветвление, т.е. образование системы побегов. Главную функцию побега – фотосинтез – осуществляют листья; стебли – преимущественно несущие органы, выполняющие механическую и проводящую функции. Главная черта, отличающая побег от корня, - его облиственность. Участок стебля, от которого отходит лист (листья) называется узел. Участки стебля между соседними узлами – междоузлия. Узлы и междоузлия повторяются вдоль оси побега. Таким образом, побег имеет метамерное строение, метамером (повторяющимся элементом) побега являются узел с листом и пазушной почкой и нижележащее междоузлие
Первый побег растения – его главный побег, или побег первого порядка. Он образуется из зародышевого побега, заканчивающегося почечкой, которая формирует все последующие метамеры главного побега. По положению эта почка – верхушечная; пока она сохраняется, данный побег способен к дальнейшему росту в длину с образованием новых метамеров. Кроме верхушечной, на побеге образуются боковые почки. У семенных растений они находятся в пазухах листьев и называются пазушными. Из боковых пазушных почек развиваются боковые побеги, и происходит ветвление, за счет которого увеличивается общая фотосинтезирующая поверхность растения. Формируется система побегов, представленная главным побегом (побегом первого порядка) и боковыми (побегами второго порядка), а при повторении ветвления – боковыми побегами третьего, четвертого и последующих порядков. Побег любого порядка имеет свою верхушечную почку и способен к нарастанию в длину. ^ Почка – это зачаточный, еще не развернувшийся побег. Внутри почки заключена меристематическая верхушка побега – его апекс (рис. 4.17). Апекс представляет собой активно работающий ростовой центр, который обеспечивает формирование всех органов и первичных тканей побега. Источником постоянного самообновления апекса являются инициальные клетки апикальной меристемы, сосредоточенные на кончике апекса. Вегетативный апекс побега, в отличие от всегда гладкого апекса корня, регулярно формирует на поверхности выступы, представляющие собой зачатки листьев. Гладким остается лишь самый кончик апекса, который называют конусом нарастания побега. Его форма сильно варьирует у разных растений и не всегда имеет вид конуса, вершинная часть апекса может быть низкой, полушаровидной, плоской или даже вогнутой. Из вегетативных почек развиваются вегетативные побеги, состоящие из стебля, листьев и почек. Такая почка состоит из меристематической зачаточной оси, оканчивающейся конусом нарастания, и зачаточных листьев разного возраста. Нижние листовые зачатки вследствие неравномерного роста загибаются внутрь и прикрывают верхние, более молодые, зачатки листьев и конус нарастания. Узлы в почке сближены, так как междоузлия еще не успели вытянуться. В пазухах листовых зачатков в почке уже могут быть заложены зачатки пазушных почек следующего порядка (рис. 4.17 ). В вегетативно-генеративных почках заложен ряд вегетативных метамеров, а конус нарастания превращен в зачаточный цветок или соцветие. Генеративные, или цветочные почки заключают в себе только зачаток соцветия или одиночного цветка, в последнем случае почку называют бутоном.
Кроме обычных, экзогенных по заложению, пазушных почек у растений часто образуются придаточные, или адвентивные почки. Они возникают не в меристематической верхушке побега, а на взрослой, уже дифференцированной части органа эндогенно, из внутренних тканей. Придаточные почки могут образоваться на стеблях (тогда они обычно расположены в междоузлиях), листьях и корнях. Придаточные почки имеют большое биологическое значение: они обеспечивают активное вегетативное возобновление и размножение тех многолетних растений, у которых они имеются. В частности, при помощи придаточных почек возобновляются и размножаются корнеотпрысковые растения (малина, осина, осот, одуванчик). Корневые отпрыски – это побеги, развившиеся из придаточных почек на корнях. Придаточные почки на листьях образуются относительно редко. Если такие почки сразу дают маленькие побеги с придаточными корешками, которые отпадают от материнского листа и вырастают в новые особи, их называют выводковыми (бриофиллум).
Направление роста побегов. Побеги, растущие вертикально, перпендикулярно поверхности земли, носят название ортотропных. Горизонтально растущие побеги называются плагиотропными. Направление роста может меняться в процессе развития побега. В зависимости от положения в пространстве различают морфологические типы побегов (рис. 4.19 ). Главный побег в большинстве случаев сохраняет ортотропный рост и остается прямостоячим. Боковые побеги могут расти в различном направлении, часто они образуют разной величины угол с материнским побегом. В процессе роста побег может изменить направление с плагиотропного на ортотропное, тогда он называется приподнимающимся, или восходящим. Побеги с сохраняющимся в течение жизни плагиотропным ростом называются стелющимися. Если они образуют в узлах придаточные корни, их называют ползучими. Ортотропный рост связан определенным образом со степенью развития механических тканей. При отсутствии хорошо развитых механических тканей в удлиненных побегах ортотропный рост невозможен. Но нередко растения, не обладающие достаточно развитым внутренним скелетом, все же растут вверх. Это достигается различными путями. Слабые побеги таких растений - лиан закручиваются вокруг какой-либо твердой опоры (вьющиеся побеги), лазят с помощью различного рода шипиков, крючков, корней - прицепок (лазящие побеги), цепляются с помощью усиков различного происхождения (цепляющиеся побеги).
Листорасположение. Листорасположение, или филлотаксис – порядок размещения листьев на оси побега. Различают несколько основных типов листорасположения (рис. 4.20 ). Спиральное, или очередное листорасположение наблюдается, когда на каждом узле расположен один лист, и основания последовательных листьев можно соединить условной спиральной линией. Двурядное листорасположение можно рассматривать как частный случай спирального. При этом на каждом узле находится один лист, охватывающий широким основанием всю или почти всю окружность оси. Мутовчатое листорасположение возникает, если на одном узле закладывается несколько листьев. Супротивное листорасположение – частный случай мутовчатого, когда на одном узле образуются два листа, точно друг против друга; чаще всего такое листорасположение бывает накрест супротивным, т.е. соседние пары листьев находятся во взаимно перпендикулярных плоскостях ( рис. 4.20).
^ Типы ветвления побега. Ветвление – это образование системы осей. Оно обеспечивает увеличение общей площади соприкосновения тела растения с воздушной средой, водой или почвой. Ветвление возникло в процессе эволюции еще до появления органов. В простейшем случае верхушка главной оси вильчато ветвится и дает начало двум осям следующего порядка. Это верхушечное, или дихотомическое ветвление. Верхушечным ветвлением обладают многие многоклеточные водоросли, а также некоторые примитивные растения, например плауны (рис. 4.21 ). Для остальных групп растений характерен более специализированный боковой тип ветвления. При этом боковые ветви закладываются ниже верхушки главной оси, не затрагивая ее способности к дальнейшему нарастанию. При таком способе потенциальные возможности ветвления и образования систем органов гораздо более обширны и биологически выгодны. Различают два типа бокового ветвления: моноподиальное и симподиальное (рис. 4.21 ). При моноподиальной системе ветвления каждая ось представляет собой моноподий, т.е. результат работы одной верхушечной меристемы. Моноподиальное ветвление характерно для большинства голосеменных и многих травянистых покрытосеменных растений. Большинство покрытосеменных растений, однако, ветвится по симподиальному типу. При симподиальном ветвлении верхушечная почка побега на определенном этапе отмирает или прекращает активный рост, но начинается усиленное развитие одной или нескольких боковых почек. Из них образуются побеги, замещающие побег, прекративший рост. Образовавшаяся при этом ось представляет собой симподий – составную ось, состоящую из осей нескольких последовательных порядков. Способность растений к симподиальному ветвлению имеет большое биологическое значение. В случае повреждения верхушечной почки рост оси будет продолжен боковыми побегами.
Наиболее часто встречающимся подземным метаморфозом побега является корневище (рис. 4.24). Корневищем принято называть долговечный подземный побег, выполняющий функции отложения запасных питательных веществ, возобновления, а иногда и вегетативного размножения. Корневище формируется у многолетних растений, как правило, не имеющих во взрослом состоянии главного корня. По положению в пространстве оно может быть горизонтальным, косым или вертикальным. Корневище обычно не несет зеленых листьев, но, будучи побегом, сохраняет метамерную структуру. Узлы выделяются либо по листовым рубцам и остаткам сухих листьев, либо по живым чешуевидным листьям, также в узлах располагаются пазушные почки. По этим признакам корневище легко отличить от корня. Как правило, на корневище формируются придаточные корни; из почек вырастают боковые ответвления корневища и надземные побеги. Формируется корневище или изначально как подземный орган (купена, вороний глаз, ландыш, черника), или сначала как надземный ассимилирующий побег, который затем погружается в почву с помощью втягивающих корней (земляника, медуница, манжетка). Корневища могут нарастать и ветвиться моноподиально (манжетка, вороний глаз) или симподиально (купена, медуница). В зависимости от длины междоузлий и интенсивности нарастания, различают длинные и короткие корневища и, соответственно, длиннокорневищные и короткокорневищные растения.
Близки к корневищам подземные столоны – недолговечные тонкие подземные побеги, несущие недоразвитые чешуевидные листья. Столоны служат для вегетативного размножения, расселения и захвата территории. Запасные питательные вещества в них не откладываются. У некоторых растений (картофель, земляная груша) к концу лета из верхушечных почек столонов образуются клубни (рис. 4.24 ). Клубень имеет шаровидную или овальную форму, стебель сильно утолщен, в нем откладываются запасные питательные вещества, листья редуцируются, в их пазухах формируются почки. Столоны отмирают и разрушаются, клубни перезимовывают, и на следующий год дают начало новым надземным побегам. Клубни не всегда развиваются на столонах. У некоторых многолетних растений клубневидно разрастается и утолщается основание главного побега (цикламен, капуста кольраби) (рис. 4.24 ). Функции клубня – запас питательных веществ, переживание неблагоприятного периода года, вегетативное возобновление и размножение.
Луковица
– это, как правило, подземный побег с
очень коротким уплощенным стеблем –
донцем
и чешуевидными мясистыми сочными
листьями, запасающими воду и растворимые
питательные вещества, главным образом
сахара. Из верхушечной и пазушных почек
луковиц вырастают надземные побеги, на
донце образуются придаточные корни
(рис. 4.24
). Таким образом, луковица – типичный
орган вегетативного возобновления и
размножения. Луковицы наиболее характерны
для растений из семейств лилейных
(лилии, тюльпаны), луковых (луки) и
амариллисовых (нарциссы,
гиацинты).
Клубнелуковица
внешне напоминает луковицу, но ее
чешуевидные листья не являются
запасающими; они сухие и пленчатые, а
запасные вещества откладываются в
утолщенной стеблевой части (шафран,
гладиолус).
 Рис.
4.24. Подземные
метаморфозы побега
: 1, 2, 3, 4 – последовательность развития
и строение клубня картофеля; 5 – клубень
цикламена; 6 – клубень кольраби; 7 –
луковички лилии тигровой; 8 – луковица
лука репчатого; 9 – луковица лилии; 10 –
участок длинного корневища пырея
ползучего.
Видоизменяться
могут не только подземные, но и надземные
побеги растений (рис.
4.25 ). Довольно
часто встречаются надземные
столоны. Это
плагиотропные недолговечные побеги,
функция которых вегетативное размножение,
расселение и захват территории. Если
столоны несут зеленые листья и участвуют
в процессе фотосинтеза, их называют
плетями
(костяника, живучка ползучая). У земляники
столоны лишены развитых зеленых листьев,
стебли у них тонкие и хрупкие, с очень
длинными междоузлиями. Такие более
высокоспециализированные для функции
вегетативного размножения столоны
называют усами.
Сочными,
мясистыми, приспособленными для
накопления воды могут быть не только
луковицы, но и надземные побеги, обычно
у растений, живущих в условиях недостатка
влаги. Водозапасающими органами могут
быть листья или стебли, иногда даже
почки. Такие сочные растения носят
название суккулентов.
Листовые суккуленты запасают воду в
тканях листьев (алоэ, агавы, толстянки,
родиола, или золотой корень). Стеблевые
суккуленты характерны для американского
семейства кактусовых и африканских
молочайных. Сочный стебель выполняет
водозапасающую и ассимилирующую функцию;
листья редуцированы или превращены в
колючки (рис.
4.25, 1). У
большинства кактусов стебли колонновидные
или шаровидные, листьев на них совсем
не образуется, но узлы хорошо заметны
по расположению пазушных побегов –
ареол,
имеющих вид бородавок или удлиненных
выростов с колючками или пучками
волосков. Превращение листьев в колючки
уменьшает испаряющую поверхность
растения и защищает от поедания животными.
Примером метаморфоза почки в суккулентный
орган – кочан
служит культурная капуста.
Рис.
4.24. Подземные
метаморфозы побега
: 1, 2, 3, 4 – последовательность развития
и строение клубня картофеля; 5 – клубень
цикламена; 6 – клубень кольраби; 7 –
луковички лилии тигровой; 8 – луковица
лука репчатого; 9 – луковица лилии; 10 –
участок длинного корневища пырея
ползучего.
Видоизменяться
могут не только подземные, но и надземные
побеги растений (рис.
4.25 ). Довольно
часто встречаются надземные
столоны. Это
плагиотропные недолговечные побеги,
функция которых вегетативное размножение,
расселение и захват территории. Если
столоны несут зеленые листья и участвуют
в процессе фотосинтеза, их называют
плетями
(костяника, живучка ползучая). У земляники
столоны лишены развитых зеленых листьев,
стебли у них тонкие и хрупкие, с очень
длинными междоузлиями. Такие более
высокоспециализированные для функции
вегетативного размножения столоны
называют усами.
Сочными,
мясистыми, приспособленными для
накопления воды могут быть не только
луковицы, но и надземные побеги, обычно
у растений, живущих в условиях недостатка
влаги. Водозапасающими органами могут
быть листья или стебли, иногда даже
почки. Такие сочные растения носят
название суккулентов.
Листовые суккуленты запасают воду в
тканях листьев (алоэ, агавы, толстянки,
родиола, или золотой корень). Стеблевые
суккуленты характерны для американского
семейства кактусовых и африканских
молочайных. Сочный стебель выполняет
водозапасающую и ассимилирующую функцию;
листья редуцированы или превращены в
колючки (рис.
4.25, 1). У
большинства кактусов стебли колонновидные
или шаровидные, листьев на них совсем
не образуется, но узлы хорошо заметны
по расположению пазушных побегов –
ареол,
имеющих вид бородавок или удлиненных
выростов с колючками или пучками
волосков. Превращение листьев в колючки
уменьшает испаряющую поверхность
растения и защищает от поедания животными.
Примером метаморфоза почки в суккулентный
орган – кочан
служит культурная капуста.
Колючки кактусов имеют листовое происхождение. Листовые колючки нередко встречаются и у несуккулентных растений (барбарис) (рис. 4.26, 1). У многих растений колючки имеют не листовое, а стеблевое происхождение. У дикой яблони, дикой груши, жостера слабительного в колючки метаморфизируются укороченные побеги, имеющие ограниченный рост и оканчивающиеся острием. Вид жесткой одревесневшей колючки они приобретают после опадения листьев. У боярышника (рис. 4.26, 3) колючки, образующиеся в пазухах листьев, совершенно безлистны с самого начала. У гледичии (рис. 4.25, 5 ) мощные разветвленные колючки образуются на стволах из спящих почек. Образование колючек любого происхождения, как правило, есть результат недостатка влаги. При выращивании многих колючих растений в искусственной влажной атмосфере они утрачивают колючки: вместо них вырастают нормальные листья (верблюжья колючка) или облиственные побеги (английский дрок).
Побеги ряда растений несут шипы. Шипы отличаются от колючек меньшими размерами, это выросты – эмергенцы – покровной ткани и тканей коры стебля (шиповник, крыжовник) (рис. 4.26, 4 ). Приспособление к недостатку влаги очень часто выражается в ранней утрате, метаморфозе или редукции листьев, теряющих основную функцию фотосинтеза. Это компенсируется тем, что роль ассимилирующего органа берет на себя стебель. Иногда такой ассимилирующий стебель безлистного побега остается внешне неизмененным (испанский дрок, верблюжья колючка) (рис. 4.25, 3). Дальнейшим шагом в указанной смене функций является формирование таких органов, как филлокладии и кладодии. Это уплощенные листоподобные стебли или целые побеги. На побегах иглицы (рис. 4.25, 4 ), в пазухах чешуевидных листьев, развиваются плоские листовидные филлокладии, имеющие, подобно листу, ограниченный рост. На филлокладиях образуются чешуевидные листья и соцветия, чего никогда не бывает на нормальных листьях, значит, филлокладий соответствует целому пазушному побегу. Мелкие, игольчатые филлокладии образуются у спаржи в пазухах чешуевидных листьев основного скелетного побега. Кладодии – это уплощенные стебли, сохраняющие, в отличие от филлокладиев, способность к длительному росту. Для некоторых растений характерно видоизменение листьев или их частей, а иногда целых побегов в усики, которые закручиваются вокруг опоры, помогая тонкому и слабому стеблю сохранять вертикальное положение. У многих бобовых в усики превращается верхняя часть перистосложного листа (горох, горошки, чина). В других случаях в усики превращаются прилистники (сассапариль). Очень характерные усики листового происхождения формируются у тыквенных, причем можно видеть все переходы от нормальных листьев к полностью метаморфизированным. Усики побегового происхождения можно наблюдать у винограда (рис. 4.25, 2), пассифлоры и ряда других растений.
Стебель
Стебель представляет собой ось побега, состоящую из узлов и междоузлий. Основные функции стебля – опорная (несущая) и проводящая. Стебель осуществляет связь между корнями и листьями. В многолетних стеблях обычно откладываются запасные питательные вещества. Молодые стебли, имеющие под эпидермой хлоренхиму, активно участвуют в фотосинтезе. Стебель обычно имеет цилиндрическую форму и характеризуется радиальной симметрией в расположении тканей. Однако в поперечном сечении он может быть не только округлым, но и угловатым – трех-, четырех- или многогранным, ребристым, бороздчатым, иногда совершенно плоским, сплюснутым, или несущим выступающие плоские ребра – крылатым
Лист
Лист – боковая структурная часть побега, выполняющая функции фотосинтеза, газообмена и транспирации. Первые листовые органы семенных растений – семядоли зародыша. Все последующие листья возникают экзогенно на апексе побега в виде бугорков или выступов – листовых зачатков, которые впоследствии развиваются в листья. Листья, как правило, имеют плоскую форму и дорсовентральное (спинно-брюшное) строение, в отличие от цилиндрических и радиально-симметричных осевых органов – стебля и корня. У семенных растений листья имеют ограниченный рост, так как, в отличие от осевых органов, лист растет за счет вставочных меристем и не имеет собственного меристематического апекса. Достигнув определенных размеров, лист до конца жизни остается без изменений. Лист, как правило, не производит никаких других органов. В редких случаях на листе могут образоваться придаточные почки и придаточные корни (бегонии, бриофиллум), но непосредственно на листе новый лист обычно не образуется. Сам же лист всегда располагается только на оси побега – стебле. Плоская пластинчатая форма листа создает наибольшую поверхность на единицу объема тканей, что способствует лучшему выполнению основной функции типичного зеленого листа – фотосинтеза. Плоская форма делает лист двусторонним. В зависимости от ориентации по отношению к верхушке побега выделяют верхнюю и нижнюю стороны листа. Дорсовентральность листа заключается в том, что в типичном случае верхняя и нижняя стороны резко отличаются по анатомическому строению, по характеру жилок, опушению и даже по окраске. На нижней стороне жилки более выпуклые, лист опушен сильнее, окраска нижней стороны листа более бледная, а иногда имеет фиолетовый или красный оттенок за счет антоцианов.
Морфология листа. Основной частью листа является листовая пластинка. Нижнюю часть листа, сочлененную со стеблем, называют основанием листа. Довольно часто между основанием и пластинкой формируется стеблеподобный цилиндрический или полукруглый в сечении черешок листа. В этом случае листья называют черешковыми, в отличие от сидячих листьев, не имеющих черешка. Роль черешка, кроме опорной и проводящей, состоит в том, что он долго сохраняет способность к вставочному росту и может регулировать положение пластинки, изгибаясь по направлению к свету. Основание листа может принимать различную форму. Иногда оно почти незаметно или имеет вид небольшого утолщения (листовая подушечка), например у кислицы. Часто основание разрастается, охватывая целиком узел и образуя трубку, называемую влагалищем листа. Образование влагалища особенно характерно для однодольных, в частности для злаков, а из двудольных – для зонтичных. Влагалища защищают вставочные меристемы, находящиеся в основании междоузлий, и пазушные почки, сидящие над узлами.
Часто основание листа дает парные боковые выросты – прилистники. Форма и размеры прилистников различны у разных растений. У древесных растений прилистники обычно имеют вид пленчатых чешуевидных образований и играют защитную роль, составляя главную часть почечных покровов. При этом они недолговечны и опадают при развертывании почек, так что на взрослом побеге у вполне развитых листьев прилистники не обнаруживаются (береза, дуб, липа, черемуха). Иногда прилистники имеют зеленую окраску и функционируют наравне с пластинкой листа как фотосинтезирующие органы (многие бобовые и розоцветные). Для всех представителей семейства гречишных характерно образование раструбов. Раструб образуется в результате слияния двух пазушных прилистников и охватывает стебель над узлом в виде короткой пленчатой трубки. Главная часть ассимилирующего листа - его пластинка. Если у листа одна пластинка, его называют простым. У сложных листьев на одном черешке с общим основанием располагаются две, три или несколько обособленных пластинок, иногда с собственными черешочками. Отдельные пластинки носят название листочков сложного листа, а общую ось, несущую листочки, называют рахисом. В зависимости от расположения листочков на рахисе различают перисто- и пальчатосложные листья. У первых листочки располагаются двумя рядами по обе стороны рахиса, продолжающего черешок. У пальчатосложных листьев рахиса нет, и листочки отходят от верхушки черешка. Частный случай сложного листа – тройчатосложный (рис. 4.52; 4.53).
Процесс формирования сложного листа напоминает ветвление, которое может идти до второго-третьего порядка, и тогда образуются дважды и трижды перистосложные листья. Если рахис заканчивается непарным листочком, лист называется непарноперистосложным, если парой листочков – парноперистосложным.
Форма пластинок простых листьев и листочков сложных листьев очень разнообразна. По форме листьев можно различать виды и роды растений в природе. Пластинка листа или листочка может быть цельной или расчлененной более или менее глубоко на лопасти, доли или сегменты, располагающиеся при этом перисто или пальчато. Различают перисто- и пальчатолопастные, перисто- и пальчатораздельные и перисто- и пальчаторассеченные листья (рис. 4.53). Встречаются дважды, трижды и многократно расчлененные листовые пластинки.
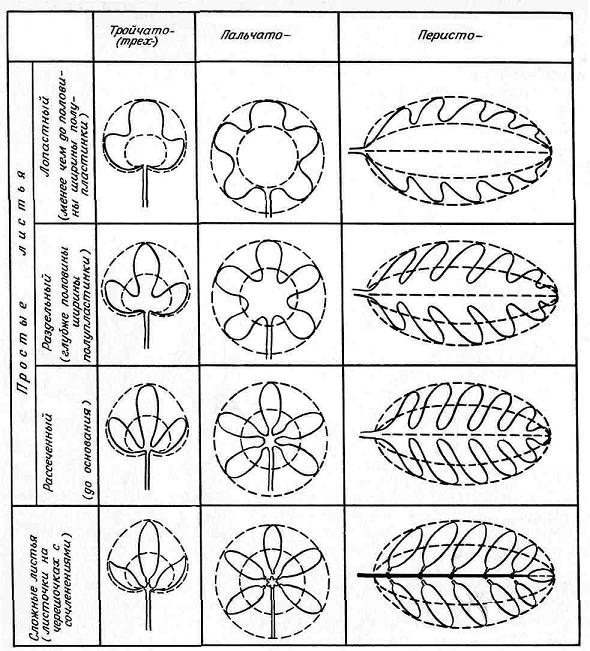 Рис.
4.53. Типы
расчленения пластинки простого
листа.
Формы
цельных листовых пластинок и расчлененных
листьев в общем очертании выделяют в
зависимости от двух параметров:
соотношения между длиной и шириной и
того, в какой части пластинки находится
ее наибольшая ширина (рис.
4.54).
Рис.
4.53. Типы
расчленения пластинки простого
листа.
Формы
цельных листовых пластинок и расчлененных
листьев в общем очертании выделяют в
зависимости от двух параметров:
соотношения между длиной и шириной и
того, в какой части пластинки находится
ее наибольшая ширина (рис.
4.54).
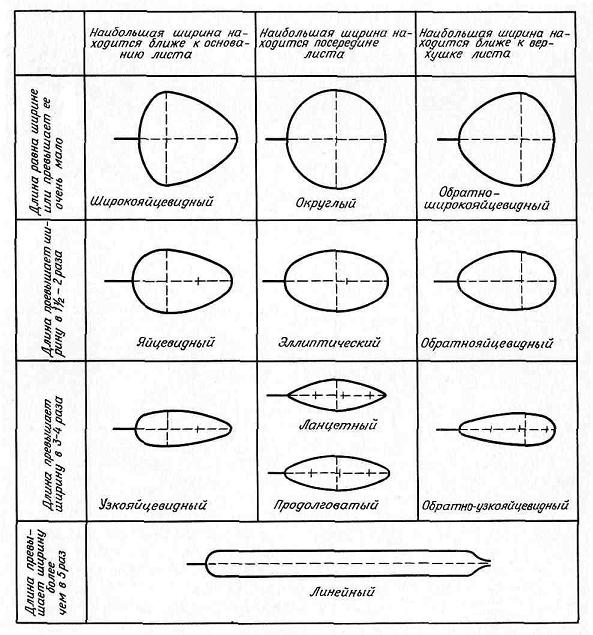 Рис.
4.54. Обобщенная
схема форм листьев.
Некоторые
особые формы листовых пластинок приведены
на рис.
4.55.
Рис.
4.54. Обобщенная
схема форм листьев.
Некоторые
особые формы листовых пластинок приведены
на рис.
4.55.
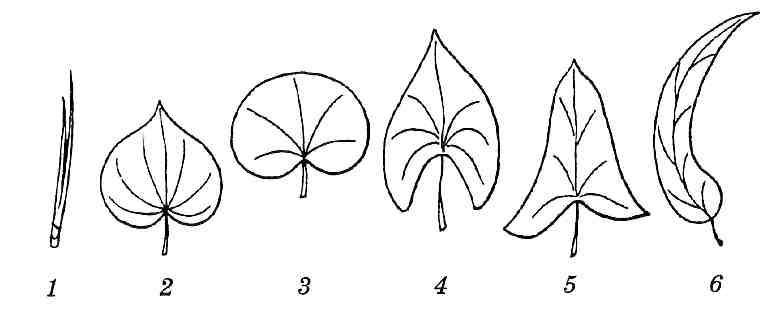 Рис.
4.55. Формы
пластинок листьев
: 1 – игольчатая; 2 – сердцевидная; 3 –
почковидная; 4 – стреловидная; 5 –
копьевидная; 6 – серповидная.
Рис.
4.55. Формы
пластинок листьев
: 1 – игольчатая; 2 – сердцевидная; 3 –
почковидная; 4 – стреловидная; 5 –
копьевидная; 6 – серповидная.
При
описании также обращают внимание на
форму верхушки, основания и края пластинки
( рис.
4.56).
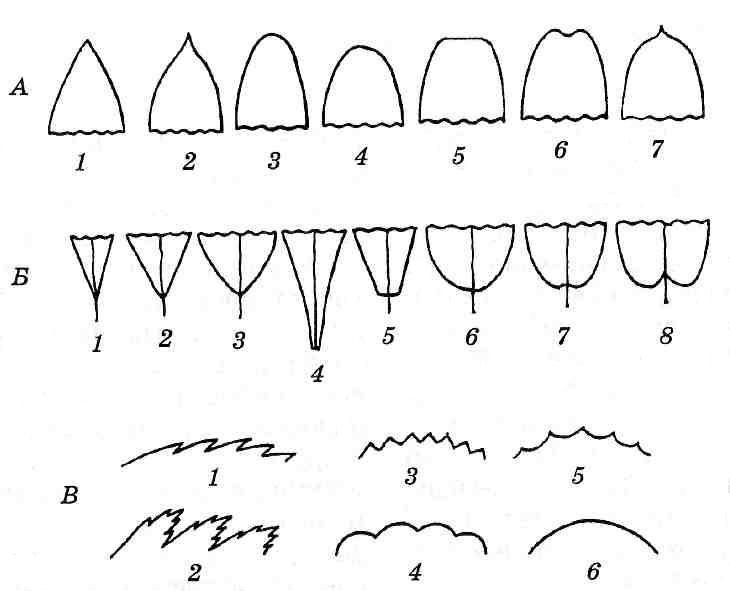 Рис.
4.56. Основные
типы верхушек, оснований и края листовых
пластинок
: А – верхушки: 1 – острая; 2 – заостренная;
3 – тупая; 4 – округлая; 5 – усеченная; 6
- выемчатая; 7 – остроконечная; Б –
основания: 1 – узкоклиновидное; 2 –
клиновидное; 3 – ширококлиновидное; 4 –
низбегающее; 5 – усеченное; 6 – округлое;
7 – выемчатое; 8 – сердцевидное; В – край
листа: 1 – пильчатый; 2 – двоякопильчатый;
3 - зубчатый; 4 – городчатый; 5 – выемчатый;
6 – цельный.
Один из
важных описательных признаков листа -
характер жилкования (рис.
4.57 ). Жилкование
– это система
проводящих пучков и сопровождающих их
тканей, посредством которых осуществляется
транспорт веществ в листе. Наиболее
примитивным является дихотомическое,
или вильчатое
жилкование, при котором жилки первого
порядка на верхушке разделяются на две
жилки второго порядка и т. д. (папоротники,
гинкго двулопастный). У большинства
хвойных в листе проходит одна или
несколько жилок, не связанных между
собой (простое
жилкование).
Рис.
4.56. Основные
типы верхушек, оснований и края листовых
пластинок
: А – верхушки: 1 – острая; 2 – заостренная;
3 – тупая; 4 – округлая; 5 – усеченная; 6
- выемчатая; 7 – остроконечная; Б –
основания: 1 – узкоклиновидное; 2 –
клиновидное; 3 – ширококлиновидное; 4 –
низбегающее; 5 – усеченное; 6 – округлое;
7 – выемчатое; 8 – сердцевидное; В – край
листа: 1 – пильчатый; 2 – двоякопильчатый;
3 - зубчатый; 4 – городчатый; 5 – выемчатый;
6 – цельный.
Один из
важных описательных признаков листа -
характер жилкования (рис.
4.57 ). Жилкование
– это система
проводящих пучков и сопровождающих их
тканей, посредством которых осуществляется
транспорт веществ в листе. Наиболее
примитивным является дихотомическое,
или вильчатое
жилкование, при котором жилки первого
порядка на верхушке разделяются на две
жилки второго порядка и т. д. (папоротники,
гинкго двулопастный). У большинства
хвойных в листе проходит одна или
несколько жилок, не связанных между
собой (простое
жилкование).
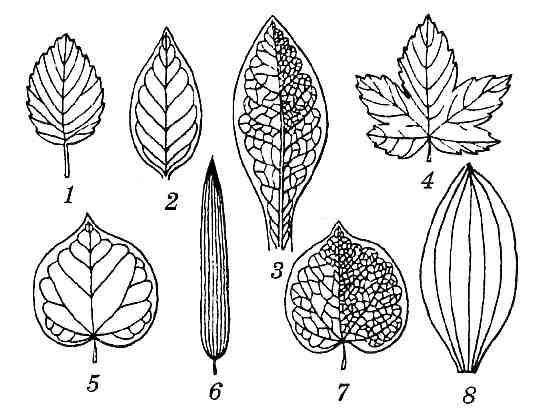 Рис.
4.57. Основные
типы жилкования листьев покрытосеменных
растений :
1 – перистокраевое; 2 – перистопетлевидное;
3 – перистосетчатое; 4 – пальчатокраевое;
5 – пальчатопетлевидное; 6 – параллельное;
7 – пальчатосетчатое; 8 – дуговидное.
У
однодольных жилки проходят вдоль листа,
не сливаясь друг с другом или отчасти
сливаясь близ верхушки. Между собой они
соединяются сетью мелких поперечных
жилок – аностомозов.
В зависимости от особенностей прохождения
жилок выделяют параллельное
и дуговидное
жилкования (рис.
4.57). При этом
лист называют параллельнонервным
и дугонервным.
У
двудольных растений имеются два основных
типа жилкования – перистое
и пальчатое.
У листьев с перистым жилкованием
(перистонервный
лист) имеется одна главная жилка,
являющаяся продолжением черешка. Она
проходит от основания пластинки к ее
верхушке. От главной жилки под углом
отходят боковые жилки первого порядка,
от них жилки второго порядка и т. д.
Пальчатонервный
лист не имеет
главной жилки. У листьев с пальчатым
жилкованием от места сочленения черешка
и пластинки отходят несколько пальцевидно
расходящихся крупных жилок первого
порядка. Если жилки первого порядка
доходят до края пластинки, выделяют
перисто-
и пальчатокраевое
(краебежное)
жилкования. Если боковые жилки образуют
петли, сливаясь не доходя до края листа,
выделяют перисто-
и пальчатопетлевидное
жилкования. Наконец, часто боковые жилки
обильно ветвятся, образуя густую сеть
аностомозирующих мелких жилок - перисто-
и пальчатосетчатое
жилкования (рис.
4.57). Мелкие
жилки образуют замкнутые участки –
ареолы.
Рис.
4.57. Основные
типы жилкования листьев покрытосеменных
растений :
1 – перистокраевое; 2 – перистопетлевидное;
3 – перистосетчатое; 4 – пальчатокраевое;
5 – пальчатопетлевидное; 6 – параллельное;
7 – пальчатосетчатое; 8 – дуговидное.
У
однодольных жилки проходят вдоль листа,
не сливаясь друг с другом или отчасти
сливаясь близ верхушки. Между собой они
соединяются сетью мелких поперечных
жилок – аностомозов.
В зависимости от особенностей прохождения
жилок выделяют параллельное
и дуговидное
жилкования (рис.
4.57). При этом
лист называют параллельнонервным
и дугонервным.
У
двудольных растений имеются два основных
типа жилкования – перистое
и пальчатое.
У листьев с перистым жилкованием
(перистонервный
лист) имеется одна главная жилка,
являющаяся продолжением черешка. Она
проходит от основания пластинки к ее
верхушке. От главной жилки под углом
отходят боковые жилки первого порядка,
от них жилки второго порядка и т. д.
Пальчатонервный
лист не имеет
главной жилки. У листьев с пальчатым
жилкованием от места сочленения черешка
и пластинки отходят несколько пальцевидно
расходящихся крупных жилок первого
порядка. Если жилки первого порядка
доходят до края пластинки, выделяют
перисто-
и пальчатокраевое
(краебежное)
жилкования. Если боковые жилки образуют
петли, сливаясь не доходя до края листа,
выделяют перисто-
и пальчатопетлевидное
жилкования. Наконец, часто боковые жилки
обильно ветвятся, образуя густую сеть
аностомозирующих мелких жилок - перисто-
и пальчатосетчатое
жилкования (рис.
4.57). Мелкие
жилки образуют замкнутые участки –
ареолы.
Конспект 5