

2 Сапин, том 1
17
нию строения человеческого тела». Наиболее известный ученик П.А. Загорского И. В. Буяльский (1789—1866), анатом и хирург, опубликовал «Анатомико-хирургические таблицы», учебник по анатомии, предложил метод бальзамирования трупов.
Выдающийся вклад в анатомическую науку (эмбриологию) внес K.M. Бэр (1792—1876) — профессор сравнительной анатомии и физиологии Петербургской медико-хирургической академии. Он открыл яйцеклетку человека и описал развитие ряда органов. В книге «Об истории развития животных» (1828—1837) он изложил учение о зародышевых листках, а также теорию рекапитуляции («повторения») основных морфологических признаков в эмбриогенезе человека.
Особое место в истории анатомии и хирургии занимают работы Н.И. Пирогова (1810—1881). Он начал свою медицинскую деятельность в стенах Московского университета и продолжал занятия анатомией и хирургией в Дерптском (ныне Тартуском) университете. По инициативе Н.И.Пирогова при Медико-хирургической академии в Петербурге был создан Анатомический институт, усовершенствована система анатомической подготовки врачей. Большой заслугой Н.И.Пирогова как анатома было открытие оригинального метода исследования тела человека по распилам замороженных трупов. Таким образом Н.И.Пирогов изучал положение органов и их взаимоотношения друг с другом и со скелетом. Результаты многолетних трудов Н.И.Пирогов обобщил в книге «Топографическая анатомия, иллюстрированная разрезами, проведенными через замороженное тело человека в трех направлениях» (1852—1859). Н.И.Пирогов изучил фасции и клетчаточные пространства в теле человека. Его труд «Хирургическая анатомия артериальных стволов и фасций» (1838) переиздавался неоднократно. Перу Н.И.Пирогова принадлежит «Полный курс прикладной анатомии человеческого тела» (1843—1848). В области анатомии Н.И.Пироговым сделано немало открытий. Именем Н.И.Пирогова названы язычный треугольник на шее, апоневроз двуглавой мышцы плеча, лимфатический узел, расположенный на глубоком кольце бедренного канала, и другие анатомические образования.
Последователь Н.И.Пирогова В.Л. Грубер (1814—1890), возглавлявший Анатомический институт в петербургской Военно-медицинской академии, создал лучший в Петербурге анатомический музей, коренным образом реорганизовал преподавание анатомии, придав ему практическую направленность. Работы В.Л.Грубера посвящены главным образом систематической и сравнительной анатомии. Они отличались точностью описания наблюдаемых фактов.
Выдающимся исследователем в области функциональной анатомии и теории физического воспитания был П.Ф. Л е с-
г а ф т (1837—1909) — автор фундаментального труда «Основы теоретической анатомии». П.Ф.Лесгафт является основоположником функциональной анатомии в России. Он описал закономерности перестройки костного вещества под влиянием мышечной тяги, сформулировал принципы развития кровеносных сосудов и их взаимоотношений в зависимости от строения и функции органов, показал значение анастомозов между артериями в кровоснабжении органов и частей тела. Работы П.Ф.Лесгафта посвящены строению и функциям суставов и мышц, общим закономерностям хода и ветвления кровеносных сосудов.
Многие открытия в области анатомии были сделаны в Киеве, входившем в то время в состав России. В.А. Б е ц (1834—1894) изучал строение мозгового вещества надпочечников, коры полушарий большого мозга, описал гигантопирамидальные невроциты (клетки Беца). Ученик Д. Н. Зернова М. А. Тихомиров (1846— 1902) изучал артерии головного мозга и варианты кровеносных сосудов. Он издал монографию «Варианты артерий и вен» (1900). Ф.А. Стефанис (1865—1917) изучал лимфатические сосуды желудка, печени, почек, организовал в Киевском университете анатомический музей.
В XX в. отечественные ученые успешно развивают функциональное и экспериментальное направления в анатомии. Разрабатываются методы научных исследований, новые перспективные научные направления. Широкое признание и распространение получил метод макро- и микроскопического исследования, успешно используются микроскопическая техника, рентгеновский и биометрический методы.
В области экспериментальной анатомии плодотворно работал основатель Ленинградской школы анатомов В.Н.Тонкое (1872—1954), в течение многих лет возглавлявший кафедру анатомии Военно-медицинской академии в Ленинграде. Его учениками были анатомы Б.А.Долго-Сабуров, Г.Ф.Иванов, А.П.Лю-бомудров, Ф.П.Маркизов, К.В.Ромодановский и др. Придавая большое значение эксперименту, В.Н.Тонков исследовал коллатеральное кровообращение, пластичность кровеносных сосудов при различных условиях существования, кровоснабжение нервов, первым (в 1896 г.) использовал рентгеновское излучение для изучения скелета. В.Н.Тонкову принадлежат также работы по эмбриологии и сравнительной анатомии. Он автор неоднократно переиздававшегося учебника по анатомии. Ученик В.Н.Тонкова и его преемник по кафедре Б.А. Долг о-С а б у-р о в (1900—1960) продолжал научное направление своего учителя. Он автор известных книг «Анастомозы и пути окольного кровообращения у человека» (1956), «Иннервация вен» (1959) и др.
М.Г. Привес (1904—2000), много лет (с 1937 по 1977) возглавлявший кафедру анатомии в I Ленинградском медицинском
2*
19
институте, известен своими работами в области ангиологии, рентгеноанатомии, бальзамирования анатомических препаратов. Он переработал и дополнил известный учебник Н.К.Лы-сенкова и В.И.Бушковича «Анатомия человека». Этим учебником студенты пользуются в течение нескольких десятилетий.
Выдающимся, широко известным в России представителем Харьковской школы анатомов был В. П. Воробьев (1876— 1937) — исследователь вегетативной нервной системы, автор методов изучения нервов. В.П.Воробьев описал нервные сплетения сердца и желудка у человека, одним из первых стал изучать иннервацию методом электростимуляции. Он создал пятитомный «Атлас анатомии человека». Ряд работ В.П.Воробьева посвящен технике приготовления музейных анатомических препаратов.
Основоположником школы отечественных лимфологов является Г.М. Иосифов (1870—1933) — профессор анатомии вначале Томского, а затем Воронежского медицинских институтов. Широко известен его труд «Лимфатическая система человека с описанием аденоидов и органов движения лимфы» (1914).
Учение об индивидуальной изменчивости и крайних формах строения тела человека и его органов разработал В.Н. Шев-к у н е н к о (1872—1952). Известны его «Курс топографической анатомии» (1935), «Атлас периферической нервной и венозной систем» (1949).
Большой вклад в анатомическую науку внес В.В. К о в а н о в (1909—1995), разработавший учение о фасциях, соединительнотканных влагалищах сосудов и нервов. Опубликованы его книги и учебники по топографической анатомии.
Кафедру анатомии Московского университета в начале XX в. (1900—1930) возглавлял ученик Д.Н.Зернова П.И. К а р у з и н (1864—1939) — организатор кафедр анатомии в ряде медицинских институтов (Астрахань, Смоленск и др.). Он автор «Руководства по пластической анатомии» (1921) и «Словаря анатомических терминов» (1928). Преемником П.И.Карузина по кафедре в Московском университете (с 1930 г. — I Московский медицинский институт) был ученик В.Н.Тонкова Г.Ф. Иванов (1893—1955) — автор книг «Хромаффинная и интерреналовая системы человека» (1930), «Нервы и органы чувств сердечно-сосудистой системы» (1945), двухтомного руководства по анатомии (1949).
Большой вклад в изучение функциональной анатомии лимфатической системы человека и животных внес ученик Г.М.Ио-сифова Д.А. Ж д а н о в (1908—1971), профессор анатомии Горьковского, затем Томского медицинских институтов, Ленинградского санитарно-гигиенического, а с 1956 — I Московского медицинского института, воспитавший многочисленных учеников. За монографию «Хирургическая анатомия грудного протока и главных лимфатических коллекторов и узлов туловища» (1945) он удостоен Государственной премии СССР.
В 1952 г. вышла его монография «Общая анатомия и физиология лимфатической системы», в 1955 г. — книга «Леонардо-да-Винчи — анатом».
Кафедрой анатомии во II Московском государственном университете (с 1930 г. — II Московский медицинский институт) до 1944 г. руководил проф. A.A. Д е ш и н (1869—1945), известный исследователь проводящих путей головного и спинного мозга. В связи с организацией во II ММИ в 1930 г. педиатрического факультета вновь созданную при нем кафедру анатомии с 1931 по 1953 г. возглавлял проф. П.П. Дьяконов (1882—1953). С 1944 г. кафедрой анатомии лечебного факультета этого института заведовал В.Н. Терновский (1888—1976) — известный историк анатомии, инициатор издания на русском языке трудов Везалия, Галена и др.
Значительный след в истории анатомической науки оставил М.Ф. Иваницкий (1895—1969), работавший в области динамической и проекционной анатомии, возглавлявший кафедру анатомии в Московском институте физической культуры, написавший оригинальный учебник по анатомии для студентов институтов физической культуры.
С 1978 по 1998 г. кафедру анатомии и спортивной морфологии в Центральном институте (академии) физической культуры возглавлял проф. Б.А. Н и к и т ю к (1933—1998), известный своими работами в области анатомической антропологии, автор ряда учебников и учебных пособий, а также таких книг, как «Конституция человека» (1991), «Очерки теории интегративной антропологии» (1995), «Биотехнологические и валеологические аспекты анатомии человека» (1997) и др.
Вопросы сравнительной анатомии, палеонтологии и антропологии разрабатывал М.А. Гремяцкий (1887—1963) — автор учебника для биологов «Анатомия человека». Широко известны исследования в области антропологии, выполненные В.В. Бунаком (1891-1979).
Заметный след в истории отечественной анатомии оставил С.С. Михайлов (1919—1993) — профессор Оренбургского, а затем (с 1964 г.) Московского стоматологического медицинского института, автор ряда монографий («Артериовенозные сонно-пещеристые аневризмы», 1965; «Иннервация интра- и экстракраниальных венозных образований», 1965, и др.). С.С.Михайлов — автор учебника «Анатомия человека» для студентов стоматологических институтов.
В середине XIX в. в медицинских вузах России появляются кафедры гистологии, отделившиеся от кафедр анатомии или физиологии. Курс гистологии и эмбриологии в петербургской Медико-хирургической академии с 1852 г. читал проф. Н.М. Якубович (1816—1879), внесший большой вклад в микроскопическую анатомию центральной нервной системы. (Кафедра гистологии и эмбриологии в Академии была открыта лишь в 1868 г.) Профессор этой кафедры М.Д. Лавдовский (1846—1902) известен своими работами в области регенерации и дегенерации нервных волокон. Он является автором (совместно с Ф.В.Овсянниковым) первого русского руководства по гистологии (1887). Сменивший М.Д.Лавдовского на этой кафедре проф. A.A. Максимов (1874—1928) был автором популярного учебника по гистологии. Он известен также своими работами в области кроветворения, изучал строение и функции соединительной ткани, крови.
В Петербургском университете вопросами гистофизиологии занимался проф. Ф.В. Овсянников (1827—1906). Его исследования были посвящены тонкому строению нервной системы и органов чувств.
Кафедру гистологии в Московском университете в 1869 г. организовал проф. А.И. Б а б у х и н (1827—1891), он же возглавил Московскую школу гистологов. А.И.Бабухин изучал гистогенез и гистофизиологию мышечной и нервной тканей, развитие и строение сетчатки глаза, осевых цилиндров нервных волокон. Ученик и преемник А.И.Бабухина по кафедре И.Ф. О г-н е в занимался проблемами строения и физиологии клеток, тканей и органов, в том числе их изменениями при воздействии внешних и внутренних факторов.
Б.И. Лаврентьев (1892—1944) — профессор Московского университета, затем II Московского медицинского института является создателем экспериментальной нейрогистологии. Он доказал наличие синаптического перерыва в вегетативных нервных узлах на путях следования нервных импульсов, разрабатывал проблемы трофики. Его учениками и последователями были профессора Н.Г.Колосов и В.Ф.Лашков (Ленинград), М.А.Барон, Т.А.Григорьева, В.В.Португалов (Москва).
Профессор I Московского медицинского института М.А. Б а-р о н (1904—1974) известен фундаментальными исследованиями серозных оболочек (брюшина, плевра и др.). Он описал их конструкцию, морфофункциональные особенности. В.Г. Елисеев (1899—1966), возглавлявший кафедру с 1953 г., после М.А.Барона, исследовал гистофизиологию крови, соединительной ткани и ее изменения при различных воздействиях. Научные результаты он обобщил в книге «Соединительная ткань» (1961). В.Г.Елисеев вместе с учениками и сотрудниками опубликовал популярный учебник «Гистология».
Ученик В.Г.Елисеева и преемник его по кафедре проф. Ю. И.Афанасьев (1927—1997) продолжил научное направление своего учителя. Большое внимание Ю.И. Афанасьев уделял методике преподавания гистологии, совершенствованию учебника гистологии, созданного В.Г. Елисеевым и выдержавшего четыре издания.
Профессор II Московского медицинского института Т. А. Григорьева (1901—1968) свои исследования нервной, соединительной, мышечной тканей обобщила в книге «Иннервация кровеносных сосудов» (1954).
Основателем Казанской школы нейрогистологов был К. А. Арнштейн (1840—1919), возглавлявший кафедру гистологии в Казанском университете с 1872 г. Широко известны его исследования периферических нервных окончаний, он описал методы выявления нервных структур с применением метиленового синего. Большой вклад в развитие нейрогистологии внесли ученики К.А.Арнштейна: А.С.Догель, Б.И.Лаврентьев, А.Н.Миславский. A.C. Догель (1852—1922) работал в Казанском, Томском (основал кафедру гистологии в 1888 г.), Петербургском университетах, изучал строение вегетативной нервной системы. В 1916 г. он основал журнал «Архив анатомии, гистологии и эмбриологии», который в 1992 г. получил новое название — «Морфология».
Ученик А.С.Догеля A.A. Заварзин (1886—1945), профессор Военно-медицинской академии в Ленинграде, — один из основоположников эволюционной гистологии. Он изучал сравнительную гистологию нервной системы, соединительной и мышечной тканей, разработал теорию тканевой эволюции, предложил классификацию тканей на основе их функций. Исследования А.А.Заварзина составили основу его фундаментальных работ: «Очерки по эволюционной гистологии нервной системы» (1941), «Очерки по эволюционной гистологии крови и соединительной ткани» (1945).
Основоположником эволюционной гистологии вместе с А.А.Заварзиным был профессор ленинградской Военно-медицинской академии Н.Г. X л о п и н (1897—1961). Он исследовал развитие и реактивность мышечной ткани, сосудистого эндотелия, нейроглии, предложил теорию дивергентного развития тканей, опубликовал монографии «Культура тканей» (1940), «Общебиологические и экспериментальные основы гистологии» (1946).
Профессор Военно-медицинской академии СИ. Щелкунов (1904—1977) успешно развивал исследования в области эволюционной и сравнительной гистологии, изучал реактивность мышечной и эпителиальной тканей, эндокарда и кровеносных сосудов. Разработал общую теорию гистогенеза (детерминации, интеграции, гетерохронности клеточных структур). Результаты его исследований обобщены в книгах «Клеточная теория и учение о тканях» (1958), «Основные принципы клеточной дифференцировки» (1977). Совместно с А.А.Заварзиным С.И.Щелкунов издал рукводство по гистологии (1954).
Вопросами эмбриологии в Казанском, а затем Ленинградском университете занимался П.Г. Светлов (1892—1974). Он установил критические периоды в эмбриогенезе, изучал влияние факторов внешней среды на развитие зародыша. Б.П. Т о-к и н (1900—1984) внес большой вклад в развитие отечественной эмбриологии. Он автор популярного учебника «Общая эмбриология», выдержавшего несколько изданий.
Во второй половине XX в. в анатомии, цитологии, гистологии и эмбриологии применяются новые, современные методы исследования. Микроскопическая и ультрамикроскопическая техника, авторадиография, рентгеновский и биометрический методы позволили получить много новых научных фактов, выявить важные закономерности. Полученные при помощи этих методов результаты исследований позволили сделать ценные для науки и практической медицины обобщения.
Вопросы для повторения и самоконтроля
Перечислите заслуги К.Галена в анатомии.
Назовите ученого, которого заслуженно считают основоположником научной описательной анатомии, расскажите о его вкладе в анатомию.
Назовите отечественные труды X—XVI вв., содержащие сведения об анатомическом строении тела человека.
Чем известен Леонардо-да-Винчи как анатом?
Перечислите известных Вам анатомов XVII—XIX вв., назовите их открытия в анатомии.
В какое время и где в России были открыты первые учебные заведения, в которых преподавалась анатомия?
В каких городах и учебных заведениях России впервые начали преподавать гистологию и эмбриологию?
Перечислите отечественных анатомов XIX в. и первой половины XX в., назовите их заслуги.
Назовите отечественных ученых, внесших существенный вклад в изучение микроскопической анатомии (гистофизиологии) органов и тканей.
ПЛОСКОСТИ и оси.
ТЕРМИНЫ ОБЩЕГО НАЗНАЧЕНИЯ
Для обозначения положения тела человека в пространстве, расположения его частей относительно друг друга в анатомии используют понятия о плоскостях и осях (рис. 1). Исходным принято считать такое положение тела, когда человек стоит, ноги вместе, ладони обращены вперед. Человек, как и другие позвоночные, построен по принципу двусторонней (билатеральной) симметрии, тело его делят на две половины — правую и левую. Границей между ними является срединная (медианная) плоскость, расположенная вертикально и ориентированная спереди назад в сагиттальном направлении (от лат. sag^tta — стрела). Эту плоскость называют также сагиттальной.
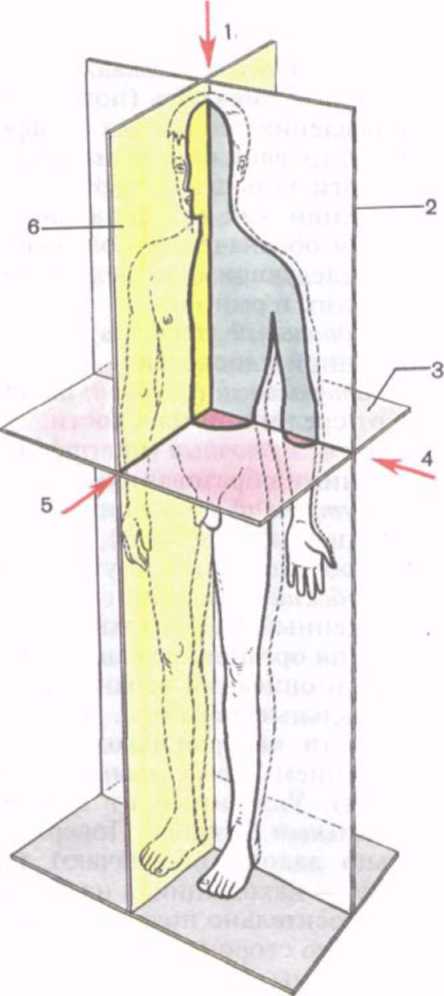
Рис. 1. Оси и плоскости, проводимые через тело человека (схема).
— вертикальная (продольная) ось;
— фронтальная плоскость; 3 — горизонтальная плоскость; 4 — поперечная ось; 5 — сагиттальная ось; 6 — сагиттальная плоскость.
Сагиттальная плоскость
отделяет правую часть тела (правый — dexter) от левой (левый — sinister). Вертикальная плоскость, ориентированная перпендикулярно к сагиттальной и отделяющая переднюю часть тела (передний — anterior) от задней (задний — posterior), называется фронтальной (от лат. fröns — лоб). Эта плоскость по своему направлению соответствует плоскости лба.
В качестве синонимов терминов «передний» и «задний» при определении положения внутренних органов можно использовать понятия «брюшной» или «вентральный» (venträlis) и «спинной» или «дорсальный» (dorsilis) соответственно.
Горизонтальная плоскость ориентирована перпендикулярно сагиттальной и фронтальной и отделяет расположенные ниже отделы тела (нижний — inferior) от вышележащих (верхний — superior).
Эти три плоскости: сагиттальная, фронтальная и горизонтальная — могут быть проведены через любую точку тела человека. Поэтому количество плоскостей может быть произвольным. Соответственно плоскостям можно выделить направления (оси), которые позволяют ориентировать органы относительно положения тела. Вертикальная ось (вертикальный — vertical is) направлена вдоль тела стоящего человека. По этой оси располагаются позвоночный столб и лежащие вдоль него органы (спинной мозг, грудная и брюшная части аорты, грудной проток, пищевод). Вертикальная ось совпадает с продольной осью (продольный — longitudinälis), которая также ориентирована вдоль тела человека независимо от его положения в пространстве или вдоль конечности (нога, рука), или вдоль органа, длинные размеры которого преобладают над другими размерами. Фронтальная (поперечная) ось (поперечный — transversus, transversalis) по направлению совпадает с фронтальной плоскостью. Эта ось ориентирована справа налево или слева направо. Сагиттальная ось (сагиттальный — sagittälis) расположена в переднезаднем направлении, как и сагиттальная плоскость.
Для обозначения положения органов и частей тела пользуются следующими определениями, входящими в список анатомических терминов:
медиальный (medialis), если орган (органы) лежит ближе к срединной плоскости;
латеральный (боковой; lateralis), если орган расположен дальше от срединной плоскости;
промежуточный (intermedius), если орган лежит между двумя соседними образованиями;
внутренний (лежащий внутри; internus) и наружный (лежащий кнаружи; externus), когда говорят об органах, расположенных соответственно внутри, в полости тела, или вне ее;
глубокий (лежащий глубже; profundus) и поверхностный (расположенный на поверхности; superficialis) для определения положения органов, лежащих на различной глубине.
При описании верхней и нижней конечностей употребляют специальные термины. Для обозначения начала конечности — той части, которая находится ближе к туловищу, пользуются определением проксимальный (ближайший к туловищу) (рго-ximälis). Удаленный от туловища отдел конечности называют дистальным (distälis). Поверхность верхней конечности относительно ладони обозначают термином ладонный (palmaris или volaris — находящийся на стороне ладони), а нижней конечности относительно подошвы — подошвенный (plantaris). Край предплечья со стороны лучевой кости называется лучевым (radialis), а со стороны локтевой кости — локтевым (ulnäris). На голени край, где располагается малоберцовая кость, называется малоберцовым (flbuläris), а противоположный край, где лежит боль-шеберцовая кость, — большеберцовым (tibiälis).
Для определения проекции границ сердца, легких, печени, плевры и других органов на поверхности тела условно проводят вертикальные линии, ориентированные вдоль тела человека. Передняя срединная линия (linea mediana anterior) проходит вдоль передней поверхности тела человека, на границе между правой и левой его половинами. Задняя срединная линия (linea mediana posterior) идет вдоль позвоночного столба, над вершинами остистых отростков позвонков. Между двумя этими линиями с каждой стороны можно провести еще несколько условных линий через анатомические образования на поверхности тела. Грудинная (око-логрудинная) линия (linea sternalis) идет по краю грудины, среднеключичная линия (linea medioclaviculdris) проходит через середину ключицы. Нередко эта линия совпадает с положением соска молочной железы, в связи с чем ее называют также сосковой линией (linea mamillaris). Передняя подмышечная линия (linea axillaris anterior) начинается от одноименной складки (plica axillaris anterior) в области подмышечной ямки и идет вдоль тела. Средняя подмышечная линия (linea axillaris media) начинается от самой глубокой точки подмышечной ямки; задняя подмышечная линия (linea axillaris posterior) — от одноименной складки (plica axillaris posterior). Лопаточная линия (linea scapularis) проходит через нижний угол лопатки, околопозвоночная линия (linea paravertebralis) — вдоль позвоночного столба через реберно-поперечные суставы (поперечные отростки позвонков).
КЛЕТКИ И ТКАНИ
Для понимания особенностей строения и функций многочисленных органов тела человека целесообразно вначале рассмотреть организацию клеток и тканей, из которых построены все эти органы и части тела. Это в первую очередь клетки и их производные, которые образуют ткани органов, систем и аппаратов органов, которые складываются в единый целостный организм.
КЛЕТКИ
Согласно современным представлениям, каждая клетка является универсальной структурно-функциональной единицей живого (рис. 2). Клетки всех живых организмов имеют сходное строение. Размножаются клетки только путем деления.
Клетка (се11и1а) — это элементарная упорядоченная единица живого. Она осуществляет функции реценции (распознавания), обмена веществ и энергии, размножения, роста и регенерации, приспособления к меняющимся условиям внутренней и внешней среды. Клетки разнообразны по своей форме, строению, химическому составу и функциям. В организме человека имеются плоские, шарообразные, овоидные, кубические, призматические, пирамидальные, звездчатые клетки. Встречаются клетки размером от нескольких микрометров (малый лимфоцит) до 200 микрометров (яйцеклетка).
От окружающей среды и соседних клеток содержимое каждой клетки отделено цитолеммой (плазмолеммой), которая обеспечивает взаимоотношения клетки с внеклеточной средой. Составными компонентами клетки, расположенными кнутри от цитолеммы, являются ядро и цитоплазма, которая состоит из гиалоплазмы и расположенных в ней органелл и включений.
 12'
12'
13<
Рис. 2. Схема ульт-рамикроскопическо го строения клетки.
1 — цитолемма (цито-плазматическая мембрана); 2 — пиноцитоз-ные пузырьки: 3 — клеточный центр (цито-центр); 4 — гиалоплаэ-ма; 5 — эндоплазмати-ческая сеть; 6 — ядро; 7 — ядрышко; 8 — пери-нукпеарное пространство; 9 — внутренний сетчатый аппарат (комплекс Гольджи); 10 — митохондрии; 1 \ — секреторные вакуоли; 12 — рибосомы; 13 — лизо-сомы; 14 — три последовательные стадии фагоцитоза.
Цитолемма (сую1ётта), или плазмолемма, — клеточная оболочка толщиной 9—10 нм. Она выполняет разделительную и защитную функции, воспринимает воздействия окружающей среды благодаря наличию рецепторов (функция рецепции). Цитолемма, выполняя обменные, транспортные функции, осуществляет перенос различных молекул (частиц) из окружающей клетку среды внутрь клетки и в обратном направлении. Процесс переноса внутрь клетки называется эндоцитозом. Эн-доцитоз подразделяют на фагоцитоз и пиноцитоз. При ф а-гоцитозе клетка захватывает и поглощает крупные частицы (частицы погибших клеток, микроорганизмы). При п и-ноцитозе цитолемма образует выпячивания, превращающиеся в пузырьки, в которые вовлекаются мелкие частицы, растворенные или взвешенные в тканевой жидкости. Пиноци-тозные пузырьки перемещают оказавшиеся в них части цы внутрь клетки.
Цитолемма участвует также в выведении веществ из клетки — экзоцитозе. Экзоцитоз осуществляется с помощью пузырьков, вакуолей, в которых выводимые из клетки вещества перемещаются вначале к цитолемме. Оболочка пузырьков сливается с цитолеммой, и их содержимое поступает во внеклеточную среду.
Рис.
3.
Схема
строения цитолеммы.
1
— липиды; 2 — гидрофобная зона
липидных молекул; 3 — белковые молекулы;
4 — полисахариды гликокаликса.
Рецепторная функция осуществляется на поверхности цитолеммы с помощью гликолипидов и гликопротеидов, которые способны распознавать химические вещества и физические факторы. Рецепторы клетки могут различать такие биологически активные вещества, как гормоны, медиаторы и др. Рецепция цитолеммы является важнейшим звеном при межклеточных взаимодействиях.
В цитолемме, являющейся полупроницаемой биологической мембраной, выделяют три слоя: наружный, промежуточный и внутренний. Наружный и внутренний слои цитолеммы, толщиной около 2,5 нм каждый, составляют электронно-плотный ли-пидный двойной слой (бислой). Между этими слоями находится электронно-светлая гидрофобная зона липидных молекул, ее толщина около 3 нм (рис. 3). В каждом монослое липидного бислоя имеются различные липиды: в наружном — цитохром, гликолипиды, углеводные цепи которых направлены кнаружи; во внутреннем монослое, обращенном к цитоплазме, — молекулы холестерина, АТФ-синтетаза. В толще цитолеммы располагаются молекулы белка. Одни из них (интегральные, или трансмембранные) проходят через всю толщу цитолеммы. Другие белки (периферические, или внешние) лежат во внутреннем или наружном монослое мембраны. Мембранные белки выполняют различные функции: одни являются рецепторами, другие — ферментами, третьи — переносчиками различных веществ, поскольку выполняют транспортные функции.
Наружная поверхность цитолеммы покрыта тонкофибриллярным слоем (от 7,5 до 200 нм) гликокаликса. Гл и кокали кс ^1усоса1ух) образован боковыми углеводными цепями гликолипидов, гликопротеидов и другими углеводными соединениями. Углеводы в виде полисахаридов образуют ветвящиеся цепочки, соединенные слипидами и белками цитолеммы.
Цитолемма на поверхности некоторых клеток образует специализированные структуры: микроворсинки, реснички, межклеточные соединения.
Микроворсинки (microvilli) длиной до 1—2 мкм и диаметром до 0,1 мкм — это покрытые цитолеммой пальцевидные выросты. В центре микроворсинки проходят пучки параллельных ак-тиновых филаментов, прикрепленных к цитолемме у верхушки микроворсинки и по бокам ее. Микроворсинки увеличивают свободную поверхность клеток. У лейкоцитов и клеток соединительной ткани микроворсинки короткие, у кишечного эпителия — длинные, причем их так много, что они образуют так называемую щеточную каемку. Благодаря актиновым филаментам микроворсинки подвижны.
Реснички и жгутики также подвижны, их движения маятнико-образные, волнообразные. Свободная поверхность реснитчатого эпителия дыхательных путей, семявыносящих канальцев, маточных труб покрыта ресничками длиной до 5—15 мкм и диаметром 0,15—0,25 мкм. В центре каждой реснички имеется осевой фила-мент (а к с о н е м а), образованный девятью соединенными между собой периферическими двойными микротрубочками, которые окружают аксонему. Начальная (проксимальная) часть микротрубочки заканчивается в виде базального тельца, расположенного в цитоплазме клетки и состоящего также из микротрубочек. Жгутики по своему строению похожи на реснички, они совершают согласованные колебательные движения благодаря скольжению микротрубочек друг относительно друга.
Цитолемма участвует в образовании межклеточных соединений.
Межклеточные соединения образуются в местах соприкосновения клеток друг с другом, они обеспечивают межклеточные взаимодействия. Такие соединения (контакты) подразделяются на простые, зубчатые и плотные. Простое соединение — это сближение цитолемм соседних клеток (межклеточное пространство) на расстояние, равное 15—20 нм. При зубчатом соединении выпячивания (зубцы) цитолеммы одной клетки заходят (вклиниваются) между зубцами другой клетки. Если выступы цитолеммы длинные, глубоко заходят между такими же выступами другой клетки, то такие соединения называют пальцевидными (интердигитации).
У специальных плотных межклеточных соединений цитолемма соседних клеток настолько сближена, что они сливаются друг с другом. При этом создается так называемая запирающая зона, непроницаемая для молекул. Если плотное соединение цитолеммы происходит на ограниченном участке, то образуется пятно слипания (десмосома). Десмосома представляет собой площадку высокой электронной плотности диаметром до 1,5 мкм, выполняющую функцию механической связи одной клетки с другой. Такие контакты чаще встречаются между эпителиальными клетками.
Встречаются также щелевидные соединения (нексусы), длина которых достигает 2—3 мкм. Цитолеммы у таких соединений отстоят друг от друга на 2—3 нм. Через такие контакты легко проходят ионы, молекулы. Поэтому нексусы называют также проводящими соединениями. Так, например, в миокарде через нексусы передается возбуждение от одних кардиомиоцитов другим.
ГИАЛОПЛАЗМА
Гиалоплазма (hyalopläsma; от греч. hyälinos — прозрачный) составляет примерно 53—55 % от общего объема цитоплазмы (cytopläsma), образуя гомогенную массу сложного состава. В ги-алоплазме присутствуют белки, полисахариды, нуклеиновые кислоты, ферменты. При участии рибосом в гиалоплазме синтезируются белки, происходят различные реакции промежуточного обмена. В гиалоплазме располагаются также органеллы, включения и клеточное ядро.
ОРГАНЕЛЛЫ КЛЕТКИ
Органеллы (organellae) являются обязательными микроструктурами для всех клеток, выполняющими определенные жизненно важные функции. Различают мембранные и немембранные органеллы. К мембранным органеллам, отграниченным от окружающей их гиалоплазмы мембранами, относятся эндоплазмати-ческая сеть, внутренний сетчатый аппарат (комплекс Гольджи), лизосомы, пероксисомы, митохондрии.
Мембранные органеллы клетки
Все мембранные органеллы построены из элементарных мембран, принцип организации которых аналогичен строению цитолемм. Цитофизиологические процессы связаны с постоянным слипанием, слиянием и разделением мембран, при этом возможны слипание и объединение только топологически одинаковых монослоев мембран. Так, наружный, обращенный к гиалоплазме слой любой мембраны органеллы идентичен внутреннему слою цитолеммы, а внутренний, обращенный в полость органеллы слой аналогичен наружному слою цитолеммы.
Эндонлазматическая сеть (reticulum endoplasmäticum) представляет собой единую непрерывную структуру, образованную системой цистерн, трубочек и уплощенных мешочков. На электронных микрофотографиях различают зернистую (шероховатую, гранулярную) и незернистую (гладкую, агранулярную) эндо-плазматическую сеть. Внешняя сторона зернистой сети покрыта рибосомами, незернистая лишена рибосом. Зернистая эндо-плазматическая сеть синтезирует (на рибосомах) и транспортирует белки. Незернистая сеть синтезирует липиды и углеводы и участвует в их обмене [например, стероидные гормоны в корковом веществе надпочечников и клетках Лейдига (сустеноцитах) яичек; гликоген — в клетках печени]. Одной из важнейших функций эндоплазматической сети является синтез мембранных белков и липидов для всех клеточных органелл.
Внутренний сетчатый аппарат, или комплекс Гольджи (apparatus reticularis internus), представляет собой совокупность мешочков, пузырьков, цистерн, трубочек, пластинок, ограниченных биологической мембраной. Элементы комплекса Гольджи соединены между собой узкими каналами. В структурах комплекса Гольджи происходят синтез и накопление полисахаридов, белково-углеводных комплексов, которые выводятся из клеток. Так образуются секреторные гранулы. Комплекс Гольджи имеется во всех клетках человека, кроме эритроцитов и роговых чешуек эпидермиса. В большинстве клеток комплекс Гольджи расположен вокруг или вблизи ядра, в экзокринных клетках — над ядром, в апикальной части клетки. Внутренняя выпуклая поверхность структур комплекса Гольджи обращена в сторону эндоплазматической сети, а внешняя, вогнутая, — к цитоплазме.
Мембраны комплекса Гольджи образованы зернистой эндоплазматической сетью и переносятся транспортными пузырьками. От внешней стороны комплекса Гольджи постоянно отпочковываются секреторные пузырьки, а мембраны его цистерн постоянно обновляются. Секреторные пузырьки поставляют мембранный материал для клеточной мембраны и гликокалик-са. Таким образом обеспечивается обновление плазматической мембраны.
Лизосомы (lysosomae) представляют собой пузырьки диаметром 0,2—0,5 мкм, содержащие около 50 видов различных гидролитических ферментов (протеазы, липазы, фосфолипазы, нук-леазы, гликозидазы, фосфатазы). Лизосомальные ферменты синтезируются на рибосомах зернистой эндоплазматической сети, откуда переносятся транспортными пузырьками в комплекс Гольджи. От пузырьков комплекса Гольджи отпочковываются первичные лизосомы. В лизосомах поддерживается кислая среда, ее рН колеблется от 3,5 до 5,0. Мембраны лизосом устойчивы к заключенным в них ферментам и предохраняют цитоплазму от их действия. Нарушение проницаемости лизосомаль-ной мембраны приводит к активации ферментов и тяжелым повреждениям клетки вплоть до ее гибели.
Во вторичных (зрелых) лизосомах (фаголизосомах) происходит переваривание биополимеров до мономеров. Последние транспортируются через лизосомальную мембрану в гиалоплаз-му клетки. Непереваренные вещества остаются в лизосоме, в результате чего лизосома превращается в так называемое остаточное тельце высокой электронной плотности.
Пероксисомы (peroxysomae) представляют собой пузырьки диаметром от 0,3 до 1,5 мкм. Они содержат окислительные фер-
менты, разрушающие перекись водорода. Пероксисомы участвуют в расщеплении аминокислот, обмене липидов, включая холестерин, пурины, в обезвреживании многих токсичных веществ. Считается, что мембраны пероксисом образуются путем отпочковывания от незернистой эндоплазматической сети, а ферменты синтезируются полирибосомами.
Митохондрии (ткоспопскй), являющиеся «энергетическими станциями клетки», участвуют в процессах клеточного дыхания и преобразования энергии в формы, доступные для использования клеткой. Их основные функции — окисление органических веществ и синтез аденозинтрифосфорной кислоты (АТФ). Митохондрии имеют вид округлых, удлиненных или палочковидных структур длиной 0,5—1,0 мкм и шириной 0,2—1,0 мкм. Количество, размеры и расположение митохондрий зависят от функции клетки, ее потребности в энергии. Много крупных митохондрий в кардиомиоцитах, мышечных волокнах диафрагмы. Они расположены группами между миофибриллами, окружены гранулами гликогена и элементами незернистой эндоплазматической сети. Митохондрии являются органеллами с двойными мембранами (толщина каждой около 7 нм). Между наружной и внутренней митохондриальными мембранами расположено межмембранное пространство шириной 10—20 нм. Внутренняя мембрана образует многочисленные складки, или кристы. Обычно кристы ориентированы поперек длинной оси митохондрии и не достигают противоположной стороны митохонд-риальной мембраны. Благодаря кристам площадь внутренней мембраны резко возрастает. Так, поверхность крист одной митохондрии гепатоцита составляет около 16 мкм . Внутри митохондрии, между кристами, находится мелкозернистый матрикс, в котором видны гранулы диаметром около 15 нм (митохондри-альные рибосомы) и тонкие нити, представляющие собой молекулы дезоксирибонуклеиновой кислоты (ДНК).
Синтезу АТФ в митохондриях предшествуют начальные этапы, происходящие в гиалоплазме. В ней (в отсутствие кислорода) сахара окисляются до пирувата (пировиноградной кислоты). Одновременно синтезируется небольшое количество АТФ. Основной синтез АТФ происходит на мембранах крист в митохондриях с участием кислорода (аэробное окисление) и ферментов, имеющихся в матриксе. При таком окислении образуется энергия для функций клетки, а также выделяются углекислота (С02) и вода (Н20). В митохондриях на собственных молекулах ДНК синтезируются молекулы информационных, транспортных и рибосомальных нуклеиновых кислот (РНК).
В матриксе митохондрий имеются также рибосомы размером до 15 нм. Однако митохондриальные нуклеиновые кислоты и рибосомы отличаются от подобных структур данной клетки. Таким образом, митохондрии имеют собственную систему, необходимую для синтеза белков и для самовоспроизведения. Уве-