

2. Біосинтез жирів.
Біосинтез жирів включає три основні процеси: біосинтез вищих жирних кислот, біосинтез гліцерину, сполучення складових частин у молекулу триацилгліцериду.
2.1. Біосинтез жирних кислот
Біосинтез жирних кислот відбувається в основному у цитоплазмі з ацетил-КоА. Основними джерелами цієї сполуки є такі процеси: розщеплення жирних кислот, розщеплення кетонних амінокислот, розщеплення вуглеводів до піровиноградної кислоти, що здатна потім перетворюватися в ацетил-КоА.
Попередником жирних кислот є малоніл-КоА, а повний синтез насичених жирних кислот каталізується синтетазним комплексом, що складається з семи ферментів.
Синтетазна система, що каталізує біосинтез жирних кислот, знаходиться в розчинній фракції цитоплазми і відповідає за наступну сумарну реакцію при біосинтезі, наприклад, пальмітинової кислоти:
СН3–СОS-KoA
+ 7НООС–СН2–СОS-KoA
+ 14НАДФН2
–> СН3–(СН2)14–СООН
+ 7СO2
+ 8HS-KoA
+ Ацетил-КоА
Малонил-КоА
+
14НАДФ
+ 6Н2O
7
У цій реакції одна молекула ацетил-КоА конденсується з сімома молекулами малоніл-КоА з утворенням однієї молекули пальмітинової кислоти. Відновлення відбувається за рахунок НАДФН2, що утворюються при фосфоглюконатному шляху перетворення глюкози.
[1]. Малоніл-КоА утворюється з цитоплазматичного ацетил-КоА та вуглекислого газу під дією ферменту ацетил-КоА-карбоксилази, коферментом якого є біотин:
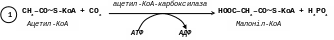
Фермент ацетил-КоА-карбоксилаза має регуляторні функції і тому лімітує весь процес синтезу жирних кислот.
[2]. Ацильні групи ацетил-КоА і малоніл-КоА переносяться на тіолові групи білка – ацилпереносника (АПБ), відповідно ацетилтрансацилазою і малонілтрансацилазою за схемою:
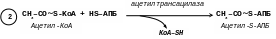

[3]. Наступним етапом є реакція конденсації, в якій ацетил-S-АПБ і малоніл-S-АПБ на поверхні ферменту реагують між собою з виділенням карбоксильної групи малоніл-S-АПБ:
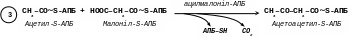
У результаті цієї реакції утворюється ацетоацетил-S-АПБ, а молекула СО2, яка виділилася, є той самий атом вуглецю, який приєднався із СО2 (див. реакцію ).
[4]. Ацетоацетил-S-АПБ відновлюється НАДФН2 з утворенням стереоізомера D--оксибутирил-S-АПБ за допомогою ферменту -кетоацил-АПБ-редуктази.
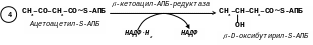
[5]. Реакція дегідратації за допомогою ферменту еноїл-АПБ-дегідратази відбувається з утворенням транс-,- або 2,3 – ненасиченого ацильного АПБ-кротоніл-S-АПБ;

[6]. Кротоніл-S-АПБ відновлюється потім кротоніл-АПБ-редуктазою до бутирил-S-АПБ. Донором водню, як і при першому відновленні, є НАДФН2.

Утворенням бутирил-S-АПБ завершується перший цикл, в якому молекула малоніл-S-АПБ приєднується до карбоксильного кінця ланцюга жирної кислоти. При цьому відщеплюється молекула АПБ, а кінцева карбоксильна група малоніл-S-АПБ відщеплюється у вигляді СО2.
Далі цикл реакцій повторюється. Допустимо, що йде синтез пальмітинової кислоти (С16). В цьому випадку утворенням бутирил-АПБ завершується лише перший з 7 циклів, в кожному з яких початком є приєднання молекули малоніл-АПБ до карбоксильного кінця ланцюга жирної кислоти, що росте. При цьому відщеплюється карбоксильна група малоніл-АПБ у вигляді СО2. Наприклад, бутиріл-АПБ, що утворився в першому циклі, взаємодіє з малоніл-АПБ:



Завершується синтез жирної кислоти відщепленням HS-АПБ від ацил-АПБ під впливом ферменту деацилази.

Отже, враховуючи, що на утворення однієї молекули малоніл-КоА з ацетіл-КоА витрачаються одна молекула АТФ і одна молекула СО2, яка потім відщеплюється, сумарне рівняння синтезу пальмітинової кислоти можна представити в такому вигляді:
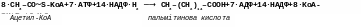
Швидкість біосинтезу жирних кислот визначається очевидно швидкістю утворення триацилгліцеридів і фосфоглідеридів тому, що вільні жирні кислоти є в клітинах у невеликих кількостях і не накопичуються.
У мітохондріях молекули насичених жирних кислот, що мають від 12 до 16 атомів вуглецю, подовжуються за рахунок послідовного приєднання ацетил-КоА.
Ненасичені жирні кислоти утворюються з насичених у клітинах за участю ферменту оксигенази.