

Тақырып: Жасанды қоректік ортада өсетін клеткалардың биологиясы
Жоспар:
1. Дедифференциялану және каллустың пайда болуы.
2. Жасушаларда атқаратын қызметіне байланысты физиологиялық және құрылымдық айырмашылықтардың пайда болуы.
3.Жасушаның белгілі тип бойынша дамуының генетикалық бағыты.
1. Дедифференциялану және каллустың пайда болуы.
Лайықты қоректік ортада кейбір клеткалар бөліну арқылы көбейіп каллус ұлпасына айналады. Каллус дедифференцияланған клеткалардан тұрады. Олай болудың себебі - әлдеқашан бөлінуін тоқтатқан, дифференцияланған клеткалар in vitro жағдайында қайтадан бөліне бастайды. Бөліну қабілетінен айырылған мұндай клеткалардың жаңадан пролиферация жолына түсіп бөліне бастауы, олардың дедифференцияланғанын көрсетеді. Басқаша айтқанда, маманданған күйін жоғалтқанын белгілейді.
Бұл процестің негізінде гендердің дифференциалды активтілігі жатады. Клеткалардың құрылымы мен қызметі гендердің активтілігіне байланысты. Организмдегі клеткалардың құрылымы меп атқаратын қызметі өзгеруіне себепші болатын, әр түрлі гендердің экспрессиясы. Яғни, клеткалардың мамандануы көптеген гендердің әрекеттесуіне байланысты. Организмнің барлық клеткаларында гендері бірдей болса да, олардың бәрі бір мезгілде әрекеттенбейді. Әдетте гендердің азғантайы ғана (5%) активті болады. Активті гендердің қатарына биологиялық түр ерекшелерін белгілейтін және клеткалық метаболизмді іске асырушы гендер кіреді. Бұдан басқа, олардың қатарына тек қана осы мүшеде, ұлпада, клеткада активті болатын гендер және белгілі бір кезеңде активтелетін гендер мен өзгерген сыртқы жағдайлардың әсерімен активтелген гендер кіреді.
Клеткаларда атқаратын қызметіне (функционалдық мамандығына) байланысты физиологиялық және құрылымдық айырмашылықтардың пайда болуын дифференциялану процесі деп атайды. Дифференциялану деген ұғым меристемалық клетканың маманданған клеткаға айналуын көрсетеді. Құрылымы және қызметі біртектес меристемалық клеткалар, әр түрлі даму жолдарына түсіп, алуан түрлі маманданған ұлпаларды түзеді. Сөйтіп, клеткалардың бір-бірінен айырмашылығы, оларда әр түрлі гендердің жұмысқа қосылуына байланысты болады. Бірақ, бүл дифференциялану процесінің іске асырылуы, клеткалық биологияның ең бір қиын, шешілмеген мәселесі болып тұр.
Жаңадан пайда болған клетканың одан әрі даму мүмкіндіктері әр қилы. Оған даму жолдарының әр қайсысына да түсуге болады. Әрбір клетканың белгілі тип бойынша дамуының генетикалық бағытын детерминация деп атайды. Ол даму физиологиясының негізі болады. Клетканың белгілі бір даму жолына түсуі ерекше белоктардың жиынтығының түзілуіне байланысты. Маманданған клеткада тек өзіне тән белоктар синтезделеді. Бұл гендердің дифференциалды активтілігінің салдары, яғни гендердің біреулері экспрессияға ілінеді, басқалары сол мезгілде репрессияланған күйде қалады. Сөйтіп клетканың фенотипі, оның қызметі белоктармен белгіленеді. Клетка қасиеттері ферменттер мен басқа белоктардың ықпалына байланысты. Ал белоктардың құрылымы иРНҚ-мен белгіленеді (3-сурет).
Сонымен, дифференцияланған кезде клетканың құрылымы мен метаболизмі өзгереді. Бірақ, биологиялық түрге тән гендер жиынтығының құрамы өзгеріске ұшырамайды. Тек қана олардың активтілік дәрежесі ғана өзгереді. Бұдан мынадай қорытынды келіп шығады: дифференциялану процесі қалай жүретінін түсіну үшін, мына негізгі мәселені шешу керек - не себептен клетканың бір типінде нақтылы бір гендер активтеледі, ал басқаларында - тіпті басқа гендер активтеледі?
Даму процесінде белоктың синтезделуі бірнеше деңгейде реттелуі мүмкін. Негізінде сондай алты деңгейі белгілі: 1) ДНҚ синтезі, яғни репликация: 2) иРНҚ синтезі, яғни транскрипция;
3) процессинг деңгейі (алғашқы иРНҚ-ның «пісіп» жетілуі, белок синтезделуіне дайын болуы); 4) и РНК-ның ядродан цитоплазмаға тасымалдануы (жетілген иРНК-лар цитоплазмаға тасымалдануы үшін сұрыпталуы); 5) трансляция деңгейі (иРНҚ-да жазылған генетикалық мәліметке сай рибосомаларда белок түзіледі);
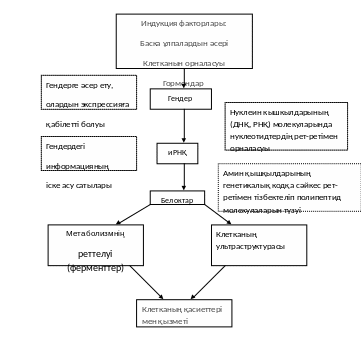
3 – сурет. Даму процесінде генетикалык информацияның іске асу жолы (Гендердің индукциясы және олардың экспрессиясы; ДНҚ →РНҚ →Белок→Клетканың құрылымы мен зат алмасуы→Клетканың генетикалық информацияға, яғни гендерге сәйкес қалыптасқан қасиеттері) (64).
6) иРНҚ-ның деградация деңгейі (цитоплазмада белок түзіліп болған соң иРНҚ-лардың ыдырауы).
ДНҚ синтезі кезіндегі реттеу. Егер ДНҚ тең емес репликацияға ұшыраса, онда клеткалардағы гендер жиынтығы әр қилы болып шығуы мүмкін. Бұдан басқа, гендер активтілігі ДНҚ-ның күйіндегі өзгерістерге де байланысты. Гендер активтілігіне әсер ететін ДНҚ өзгерісінің үш тұрақты түрі белгіленген. 1) Хромосомада гендердің өзара тұрған орындарының өзгеруі олардың функциясына ықпал етеді. Транслокация немесе инверсия кезінде ген өз орнын ауыстырып, ДНҚ, молекуласының басқа бір жеріне келіп енеді. Бұл гендердің активтілігінің төмендеуіне немесе кенет кушеюіне әкеп соғады. Орнын ауыстыратын гендерді «секіргіш гендер», транспозондар немесе мобильді (жылжымалы) дисперленген генетикалық элементтер деп атайды. Олар бір хромосомадан екіншісіне көшіп гендердің мутацияларына себепші болады немесе бұрын тежеліп тұрған гендерді активтейді. Транспозондарды Барбара Мак - Клинток 1942 жылы ашты және сол ғылыми жімістігі үшін ол Нобель сыйлығын алды. 2) Амплификация (қандай болмасын ген санының көбеюі) арқасында ген экспрессиясында өзгеріс пайда болуы мумкін. Көбінесе амплификацияға рибосомалық РНҚ-ның гендері ұшырайды. Бірақ клетканың дифференциялануына гендер амплификациясының маңызы әлі зерттелмеген. 3) Ген құрамында сапалық өзгерістер өтуі мүмкін (мысалы әр турлі мутациялар).
Белок синтезінің транскрипция деңгейінде реттелуін прокариоттарда Жакоб пен Моно зерттеген. Ал эукариоттарда гендердің активтілігінің реттелуін зерттеу өте қиын, себебі олардың геномдары өте үлкен және күрделі. Бәлки, эукариоттарда транскрипция сатысында гендердің дифференциалды активтілігі, олардың бүркеліп қалуы немесе ашылуы арқылы реттеледі. Р. Бриттен мен Э. Дэвидсон жоғары сатыдағы организмдерде гендердің реттелуін жете зерттеп өз гипотезасын ұсынды. Ядролық РНҚ – да нуклеотидтердің бірегей жүйеліліктерінің цитоплазмалық иРНҚ – сынан едәуір артық болғаны олардың назарын аударды. Олардың болжамы бойынша, гендердің экспрессиясы бастаушы иРНҚ процессингке ұшырап цитоплазмаға шығуымен реттеледі.
Р. Бриттен мен Э. Дэвидсон терминдері бойынша цитоплазмаға өтетін әр түрлі иРНҚ, тРНҚ, рРНҚ құрылымын белгілейтін ДНҚ тізбегі продюсер гені деп аталады (4-сурет). Әрбір продюсер геннің қызметі, ДНҚ молекуласында соған көршілес орналасқан тізбегімен реттеледі. Ондай нуклеотидтер тізбегі рецептор гені деп аталады. Рецептор гені РНҚ – ньң белгілі бір түрімен – активаторлық РНҚ – мен әрекеттесу арқылы активтелінеді. Активаторлық РНҚ-ның синтезі интегратор гені деген ДНҚ – ның тізбегінде өтеді. Активаторлық РНҚ бірнеше рецептор гендерімен әрекеттеседі. Сонымен бір интегратор гені бір топ түрлі ферменттердің синтезін қозғауы мүмкін. Ал интегратор геннің қызметін көрші сенсорлық гені бақылайды. Сенсоролық ген дегендері ол сыртқы факторлар, яғни индукциялық факторлары әсер ететін ДНҚ-ның бір тізбегі. Продюсер генге әр түрлі рецептор гендері сәйкес болуы мүмкін. Оның нәтижесінде бір продюсер гені әр түрлі интегратор гендер мен сенсорлық гендер арқылы алуан индукторлармен әрекеттеседі. Осының арқасында бір ферменттің түзілуі әр түрлі қоздырғьш заттармен индукциялануы түсінікті болды. Ал бір сенсорлық генге бірнеше интегратор гендері сәйкес болу арқасында белок синтезінің реттелуі одан әрі күрделенеді. Сонымен, Р. Бриттен мен Э. Дэвидсонның гендердің экспрессиясын реттеу моделі бір индукциялық фактордың көптеген алуан түрлі реакцияларды қоздыруын және біркелкі реакциялардың әр түрлі индукциялық факторлармен қоздырылуын айқын көрсетеді.
Ядродан цитоплазмаға өткен ерекше иРНҚ-ның трансляциясы лезде басталмайды. Эукариоттар эволюциясындағы маңызды ерекшелік - олардың иРНҚ-дары информосомалармен тығыз байланыста болуында. Информосомалар рибонуклеопротеид-терден тұрады, белок синтезінің реттелуін қамтамасыз етеді.
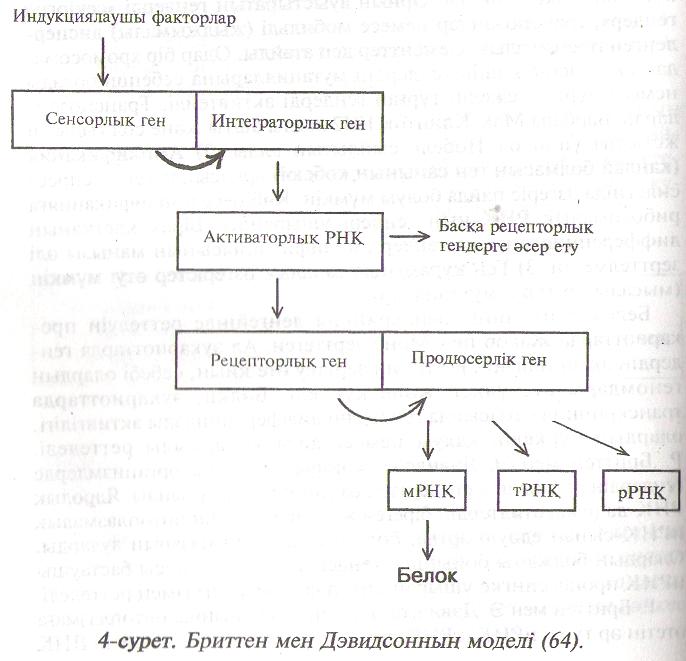
4-сурет. Бриттен мен Дэвңцсоннын моделі .
Сонымен, өсімдіктердің әр түрлі бөліктерінде немесе тіршілік циклінің кезеңдерінде өтетін дифференциялану процестері гендер экспрессиясының ерекшеліктеріне негізделген. Гендер экспрессиясының барлық тізбектеле жүретін процестерін (гендегі информацияны көшіруден, немесе транскрипциядан бастап активті белок түзілгенше, яғни трансляцияға шейін) бақылайтын механизмдері және ген экспрессиясы мен репрессиясына әсер етудің мәні мен ықпал ету жолы әлі белгісіз. Өсімдіктердің нормалы клеткалық дифференциялану процесінде ешбір ген жоғалмайды немесе біржола тежелмейді. Оны дәлелдейтін өсімдік клеткалардың тотипотенттік қасиеті, яғни жалғыз сомалық клетканың регенерация арқылы тұтас организмді түзу қабілеті. Өсімдік клеткасының дифференциациясы - қайтымды процесс болып келеді. Меристемалық клетка дифференциялану арқылы маманданған клеткаға айналады. Ал in vitro жағдайында сондай маманданған клеткалар қайтадан бөліне бастайды. Сонан соң дифференциялану жолына қайта түсіп, яғни редифференцияланып, маманданған күйге келеді. Тіпті микроспоралар сияқгы жоғары маманданған клеткалардың өздері лайықты жағдайда бөліне бастайды да қайтадан дифференциялану арқылы тұтас өсімдікті түзеді. Сөйтіп, жетілген өсімдік клеткалары бөліну қабілетін сақтайды, ал қолайлы жағдайда жаңа даму жолына ауысады. Дегенмен, сол процестерде молекулалық денгейінде өтетін құбылыстар әзірше белгісіз болып тұр.
Каллус экспланттың әр түрлі клеткаларынан пайда болады. Оларда өтетін дифференциялану процесі де бірдей болмайды. Сондықтан, олардан түзілген каллус клеткалары да бір-біріне ұқсамайды. Дифференциялану мен каллустың пайда болуының механизмдерін зерттеу үшін ең қолайлысы темекі сабағының өзекті паренхимасының клеткалары. Себебі, олардың бөлініп каллусқа айналуы ауксин мен цитокинің әсеріне тікелей байланысты. Клеткалар дедифференцияланып каллусқа айналу үшін қоректік ортада міндетті түрде фитогормондар болуы керек. Маманданған клетканың дедифференциялануының ең алғашқы кезеңі - оның бөлінуі. Р.Г. Бутенко көрсеткендей, темекінің паренхима клеткаларында in vitro өтетін дедифференциялану процесі қордағы заттарды пайдалануынан және кейбір арнайы органеллалардың бүлінуінен басталады. Дедифференциялану басталған соң 6-12 сағаттан кейін клетка құрылымында мынадай ерекше өзгерістер байқалған: клетка қабығы жұмсарған және ісінген, рибосомалар саны, эндоплазмалық тордың, Гольджи аппараты элементтерінің саны көбейген, ядрошықтардың саны көбейген және көлемі өскен. Тасымалдаушы РНҚ-ның және рибосомалық РНҚ-ның синтезі күшейген, ДНҚ-ның синтезі басталған, жаңадан синтезделген белоктар көбейген. Алғашқы ұлпада кездеспсген ерекше белоктар-антигендер пайда болады, тіпті меристема ұлпасына тән белок анықталған. Барлық атап өткен шегерістер тек қана ауксин және цитокининнің қатысуымен жүзеге асқан. Одан кейін 48-72 сағат өткен соң клеткалар бөліне бастаған. Қоректік ортаға тек қана ауксин қосылғанда, РНҚ, мен ДНҚ-ның синтезі күшейсе де, бірақ клеткалар бөлінбеген. Төрт күннен кейін клеткалар созылып өсе бастаған. Ал қоректік ортада тек цитокинин болған кезде нуклеин қышқылдарының синтезі өзгермеген. Сөйтіп, клетка бөліну үшін екі фитогормон да (ауксин және цитокинин) қажет екендігі дәлелденді.
Р.Г. Бутенко болжағандай, бұл процесс былай өтеді. Ауксиннің ықпалымен клетка мембранасы плазмалемманың өткізгіштігі артады және қасиеттері өзгереді. Одан кейін бұрыннан бар иРНҚ,-ларда немесе жаңадан синтезделген иРНҚ-ларында цитоплазмалық белоктардың синтезі күшейеді. Бұл белоктар цитоплазмадан ядроға өтіп гистондармен ерекше байланысады. Соньң арқасында рибосомалық және тасымалдаушы РНҚ-ларды синтездейтін гендер репрессияланған жағдайдан активті күйге көшеді. РНҚ мөлшері клеткада белгілі бір деңгейге жеткен соң ғана ДНҚ-ның репликациясы басталады. Ал кинетин жоқ кезде мұнан былай митоздық циклдің өтуі тежеледі. Клеткаға митозды бастауға және бөлінуді аяқтауға қажет арнайы РНҚ мен белоктардың синтезін тек цитокинин бақылайды. Клетка бөліну үшін митоз басталмай тұрып өзінің иРНҚ-сында синтезделетін ерекше бір белоктың болуы жануар клеткасында көрсетілген. Сонымен, қандай да болмасын клетканың каллус клеткасына айналу алдында әр қилы биохимиялы және структуралық терең өзгерістер өтеді.
Клетка циклі 5-ші суретте көрсетілген. Бөліну (митоз және цитокинез) нәтижесінде пайда болған жаңа клетканың ″тағдыры" екі жолмен шешілуі мүмкін: 1) клетка бөліну циклінен шығып дифференцияланған күйге біртіндеп жетуі мүмкін; 2) клетка қайтадан жаңа бөліну циклін бастап интерфаза кезеңіне түседі. Интерфаза G1 , сатысынан басталады. Бұл кезеңде биосинтез ироцестерінің қарқыны артады, мол мөлшерде қоректік заттар пайда болады, мембраналар түзіледі, түрлі органеллалар қалыптасады. Ѕ сатысында ДНҚ-ның синтезі өтеді. ДНҚ-ның мөлшері екі есе өсіп, хромосомалар саны да толығымен екі еселенген соң Ѕ сатысы аяқталады. Одан кейін клетка G2 сатысына өтеді. Бұл кезеңде клетка толығымен дамып жетіледі де қайта бөлінуге қабілетті болады. Митоз басталғанда хромосома жіпшелерінің шиыршықталып бұралуының салдарынан хромосомалар қысқарып, жуандайды. Ядро қабығы бұзылады, ал хромосомалар қозғалып орнын өзгертеді. Хромосомалар промотидтерге жіктеліп, хроматин жіпшелері клетканың полюстеріне қарай созылып, аяғында екі ядро пайда болады. Сонан соң цитокинез өтеді де цитоплазма екіге бөлінеді. Әрқайсында өз ядросы бар екі жаңа клетка пайда болады. Жас клеткалар созылу кезеңіне өтеді. Вакуольге судың көп енуіне байланысты клетка көлемі ұлғайып, клетка қабығы созылады. Сонымен қатар белоктар, нуклеин қышқылдары, көмірсулар, майлар т.с.с. заттар жаңадан пайда болады, клетка цитоплазмасы мен қабығының компоненттері түзіледі. Созылу кезеңінен кейін клетка дифференциялануға көшеді, яғни оның мамандануы басталады. 5-ші суретте көрсетілгендей, клеткалар дифферен-циялануға циклдің G1 , G2 сатыларынан тыныштық кезеңдерінен (R1 R2,) өткеннен кейін ғана шығады. Дифференцияланған клеткалар бөлінуіне бет алғанда, яғни дедифференцияланғанда, клеткалық цикліне осы G1 G2 сатылары арқылы өтеді.
Сөйтіп, әрбір жас клетканың алдында үш жол ашылады. I) Клетка эмбриональды түрінде қалып, қайтадан клеткалық цикліне түседі. 2) Бөлінбей тыныштық күйінде циклден тыс қалады (G0 ). 3) Компетенцияға иеленіп біртіндеп детерминация салдарынан дифференцияланады, яғни маманданады.
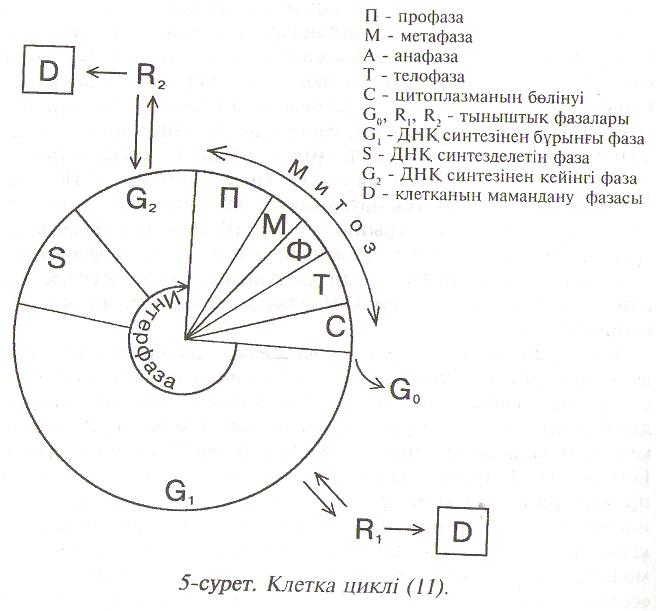
5-сурет. Клетка циклі (11).
Клетканың индуктор ықпалын қабылдауға қабілеттілігін және соның салдарынан даму жолын өзгертуге мүмкіншілік алуын компетенция деп атайды. Индуктор, яғни қоздырғыш ретінде әр түрлі факторлар болуы мүмкін: гормондар, көрші клеткалардың метаболиттері, электрофизиологиялық сигналдар т.с.с. Клетканың белгілі бір даму жолына түсуіне дайынд ылығы, яғни белгілі бір тұқым қуалаушылық қасиетін жүзеге асыруы детерминация деп аталады. Детерминация арқасында клетка белгілі бір даму жолына түскен соң, оның басқа жолдармен даму мүмкіншіліктері шектеледі. Егер клетка әуел бастан компетентті болса, оның детерминациясы бөлінген со лезде басталады. Ол тез маманданып белгілі бір ұлпаның клеткасына айналады.
Ең алғашқы каллустың пайда болуы оңай емес. Себебі, әр түрлерге жататын өсімдіктер, тіпті бір ғана өсімдіктін әр алуан ұлпалары дедифференцияланып, олар каллуска айналу үшін әр түрлі қоректік ортаны қажет етеді. Әсіресе, бұл гормондардың мөлшері мен олардың өзара қарым-қатынасына байланысты. Кейбір өсімдіктердің ұлпалары ұзақ өсіргенде өздері ауксин мен аитокининді мол синтездей бастайды, яғни гормондар жөнінде прототрофты (өз-өздерін қамтамасыз ететін) болып кетеді. Мұндай каллустарды қалыптасқан каллустар деп атайды, Ондай штамдардың пайда болуы әзірше түсініксіз. Мүмкін, олар мутация немесе гендердің дерепрессиясы нәтижесінде пайда болады. Сондай-ақ, ісік клеткалары да өсу үшін гормондарды қажет етпейді.
1. Дифференциялану және дедифференциялану деген не?
2. Компетенция мен детерминация деген не?
3. Гендер активтілігі қалай реттеледі?
4. Каллус қалай пайда болады?
5. клеткаларды жасанды ортада өсіргенде өзгергіштіктің қандай түрлері байқалады?
6. Іn vitro жағдайында өсетін клеткалар неліктен әртекті болады?
7. Іn vitrо жағдайында клеткалардың өсу заңдылығын қалай бейнелеуге болады?
4 Дәріс